

Sustained B cell depletion by CD19-targeted CAR T cells is a highly effective treatment for murine lupus
- PMID: 30842314
- PMCID: PMC8201923
- DOI: 10.1126/scitranslmed.aav1648
Sustained B cell depletion by CD19-targeted CAR T cells is a highly effective treatment for murine lupus
Abstract
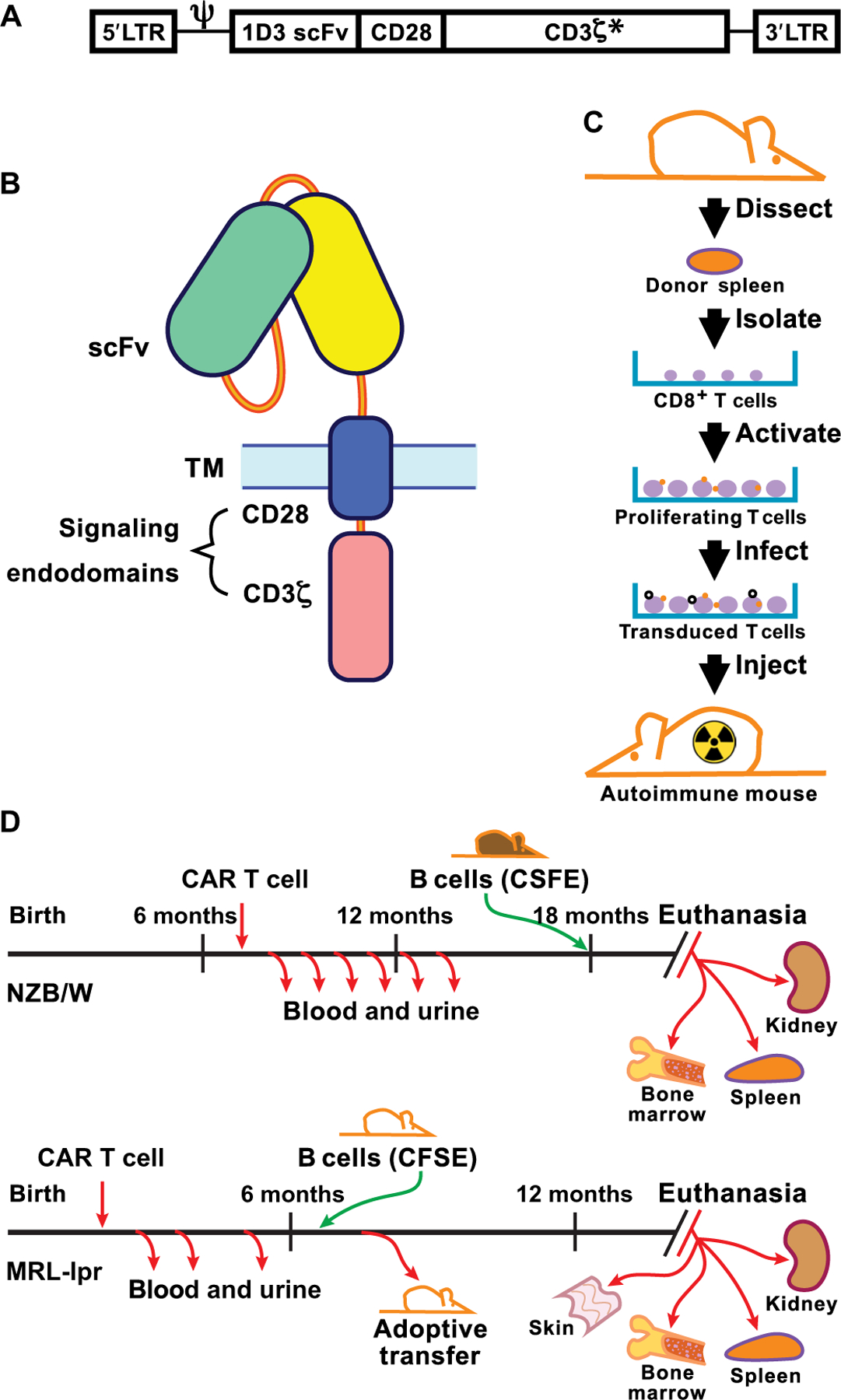
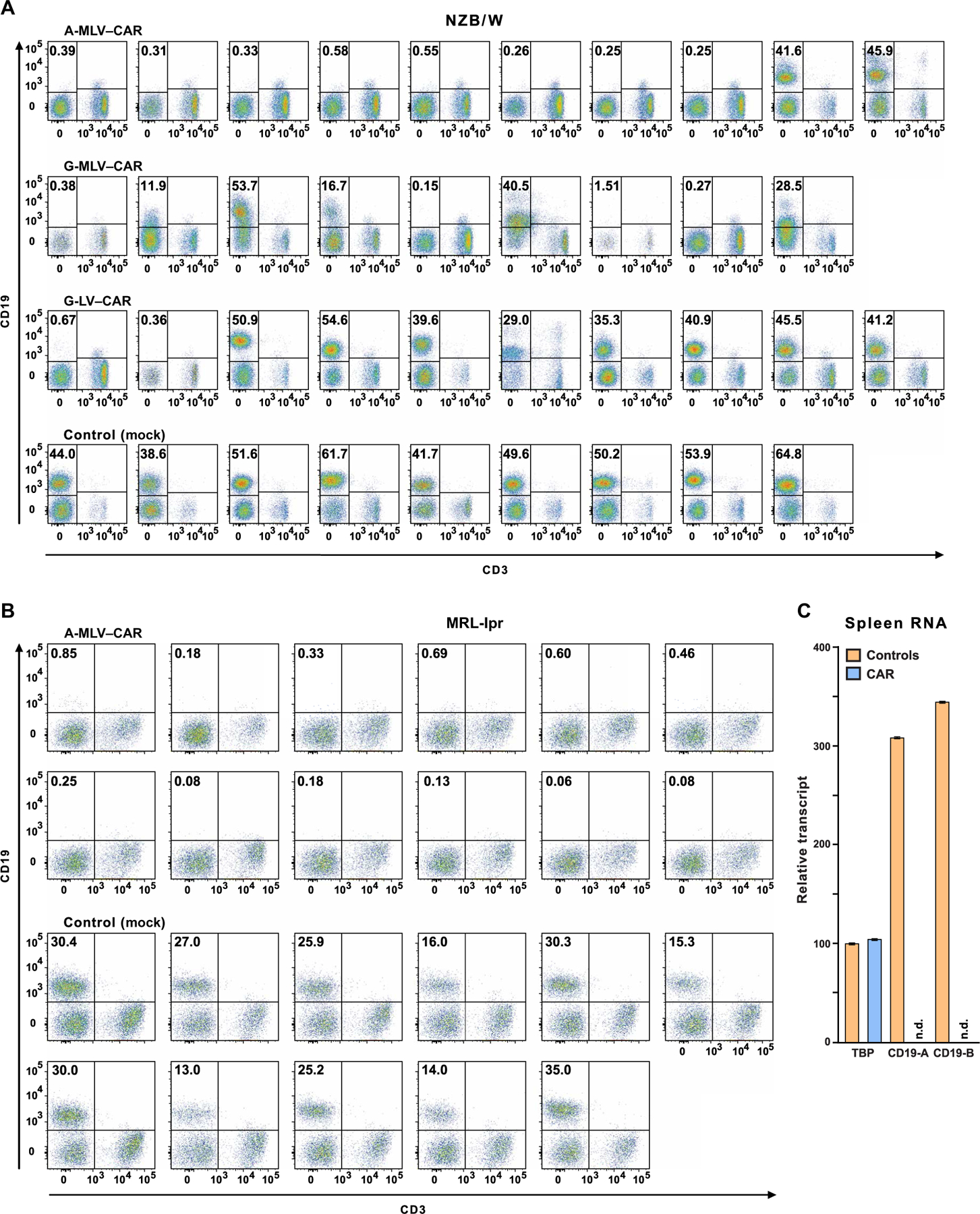
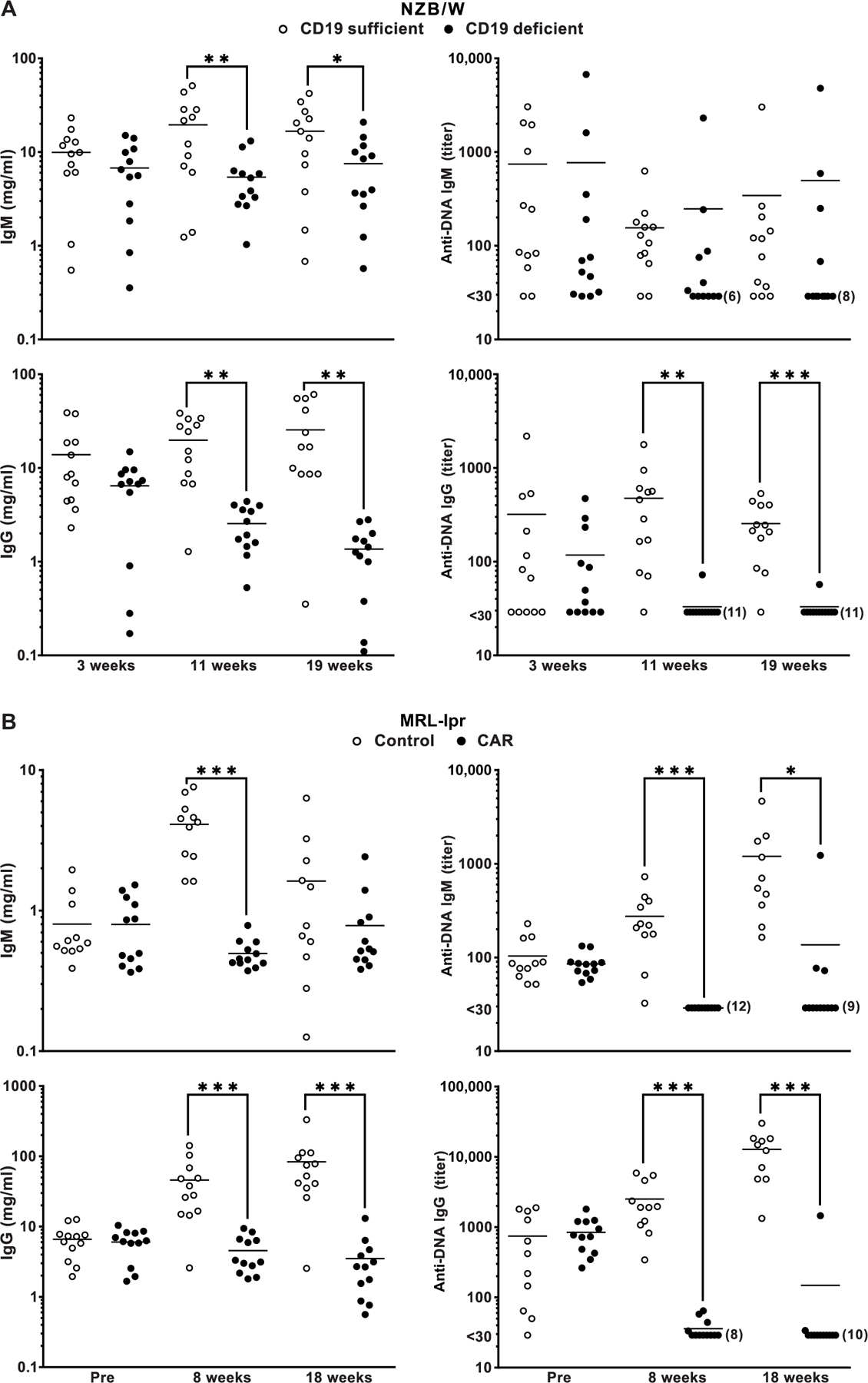
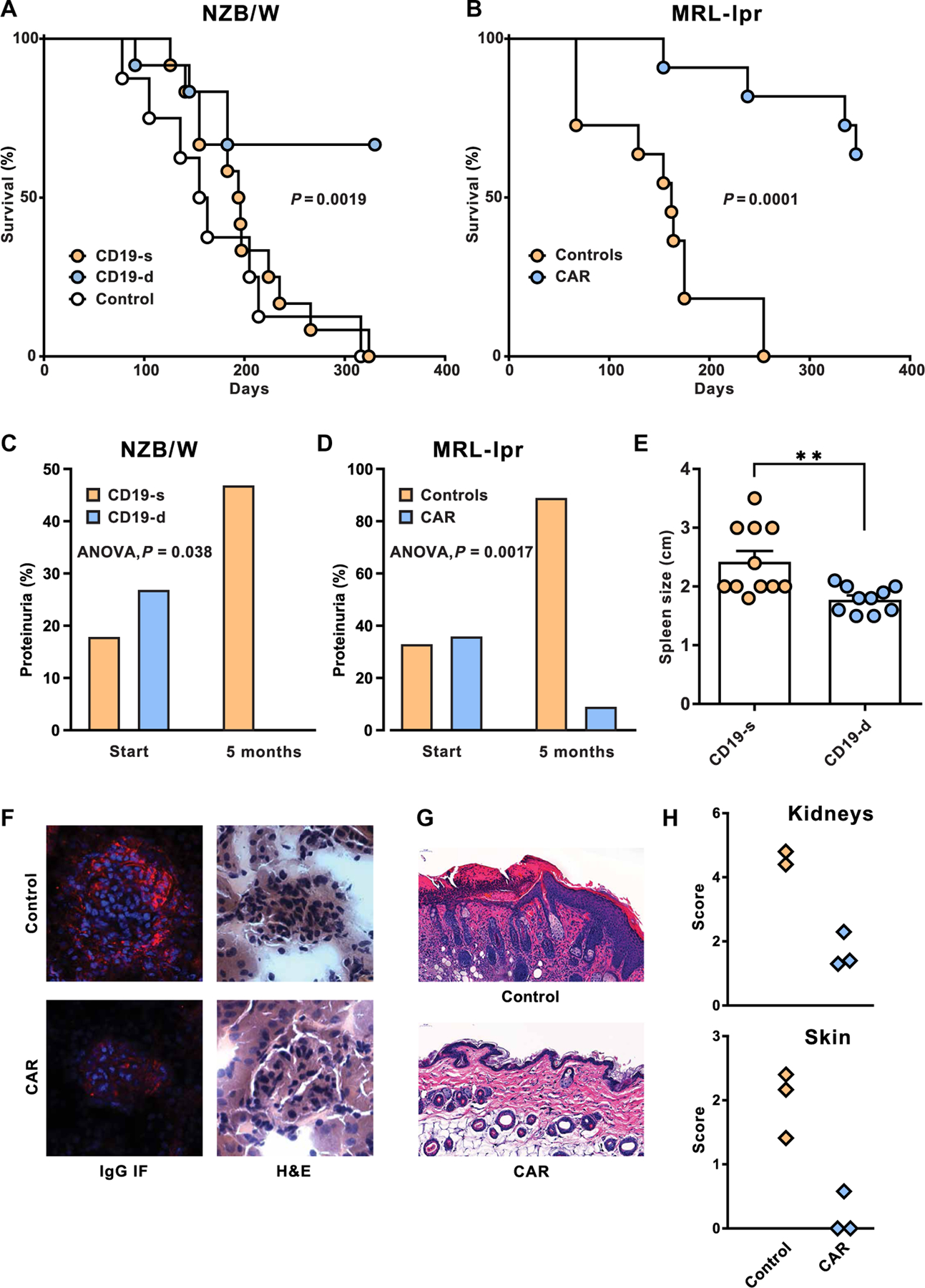
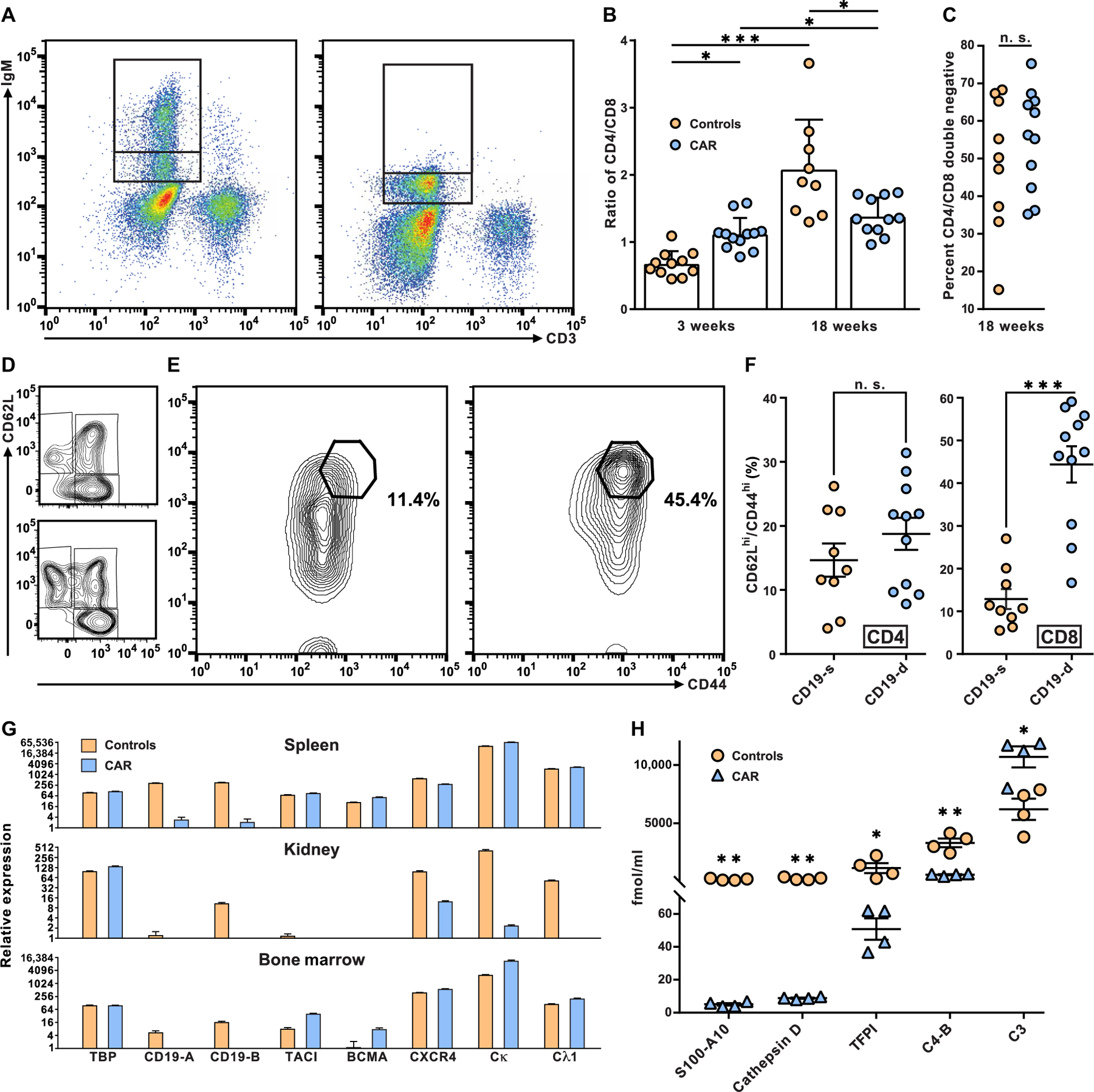
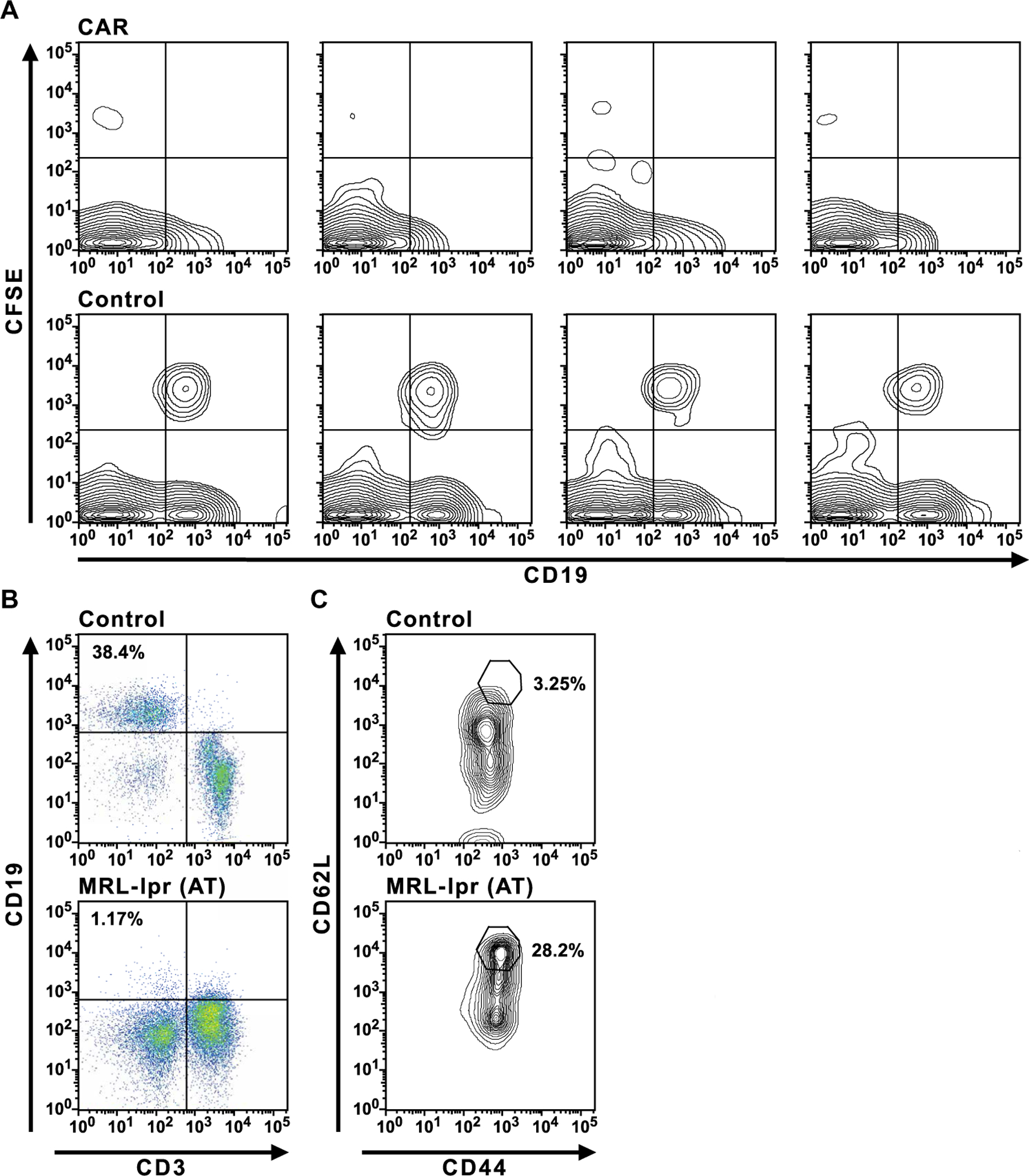
The failure of anti-CD20 antibody (Rituximab) as therapy for lupus may be attributed to the transient and incomplete B cell depletion achieved in clinical trials. Here, using an alternative approach, we report that complete and sustained CD19+ B cell depletion is a highly effective therapy in lupus models. CD8+ T cells expressing CD19-targeted chimeric antigen receptors (CARs) persistently depleted CD19+ B cells, eliminated autoantibody production, reversed disease manifestations in target organs, and extended life spans well beyond normal in the (NZB × NZW) F1 and MRL fas/fas mouse models of lupus. CAR T cells were active for 1 year in vivo and were enriched in the CD44+CD62L+ T cell subset. Adoptively transferred splenic T cells from CAR T cell-treated mice depleted CD19+ B cells and reduced disease in naive autoimmune mice, indicating that disease control was cell-mediated. Sustained B cell depletion with CD19-targeted CAR T cell immunotherapy is a stable and effective strategy to treat murine lupus, and its effectiveness should be explored in clinical trials for lupus.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Repurposing CAR T cells to treat SLE.Nat Rev Nephrol. 2019 Jun;15(6):319. doi: 10.1038/s41581-019-0144-5. Nat Rev Nephrol. 2019. PMID: 30911171 No abstract available.
-
CAR T cells drive out B cells in SLE.Nat Rev Rheumatol. 2019 May;15(5):249. doi: 10.1038/s41584-019-0214-x. Nat Rev Rheumatol. 2019. PMID: 30948844 No abstract available.
-
Déjà Vu But New: Using T Cells to Deplete B Cells to Treat Lupus.Am J Kidney Dis. 2019 Nov;74(5):708-710. doi: 10.1053/j.ajkd.2019.04.009. Epub 2019 May 30. Am J Kidney Dis. 2019. PMID: 31155322 No abstract available.
References
-
- Rahman A, Isenberg DA, Systemic lupus erythematosus. N. Engl. J. Med 358, 929–939 (2008). - PubMed
-
- Gallagher S, Yusuf I, McCaughtry TM, Turman S, Sun H, Kolbeck R, Herbst R, Wang Y, MEDI-551 treatment effectively depletes B cells and reduces serum titers of autoantibodies in mice transgenic for Sle1 and human CD19. Arthritis Rheumatol 68, 965–976 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
