

Local unfolding of the HSP27 monomer regulates chaperone activity
- PMID: 30842409
- PMCID: PMC6403371
- DOI: 10.1038/s41467-019-08557-8
Local unfolding of the HSP27 monomer regulates chaperone activity
Abstract
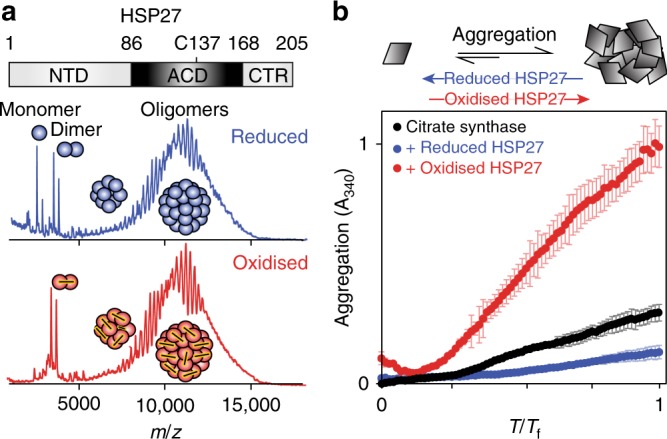
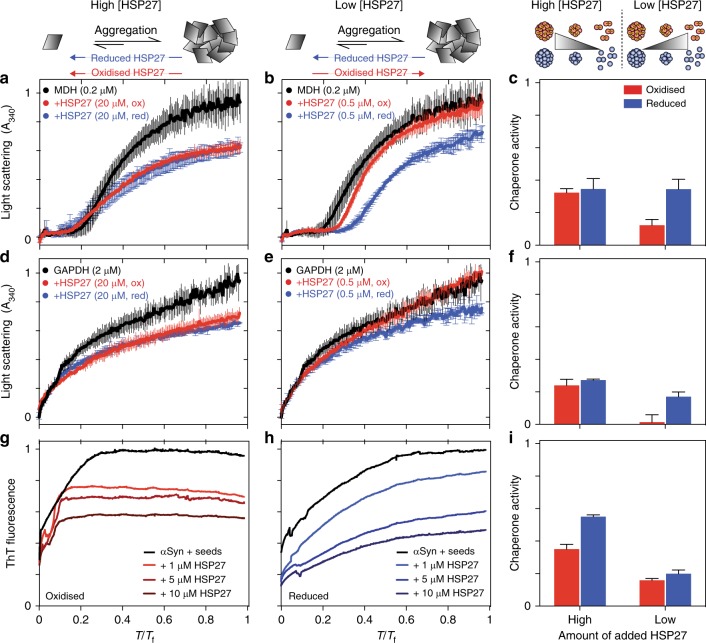
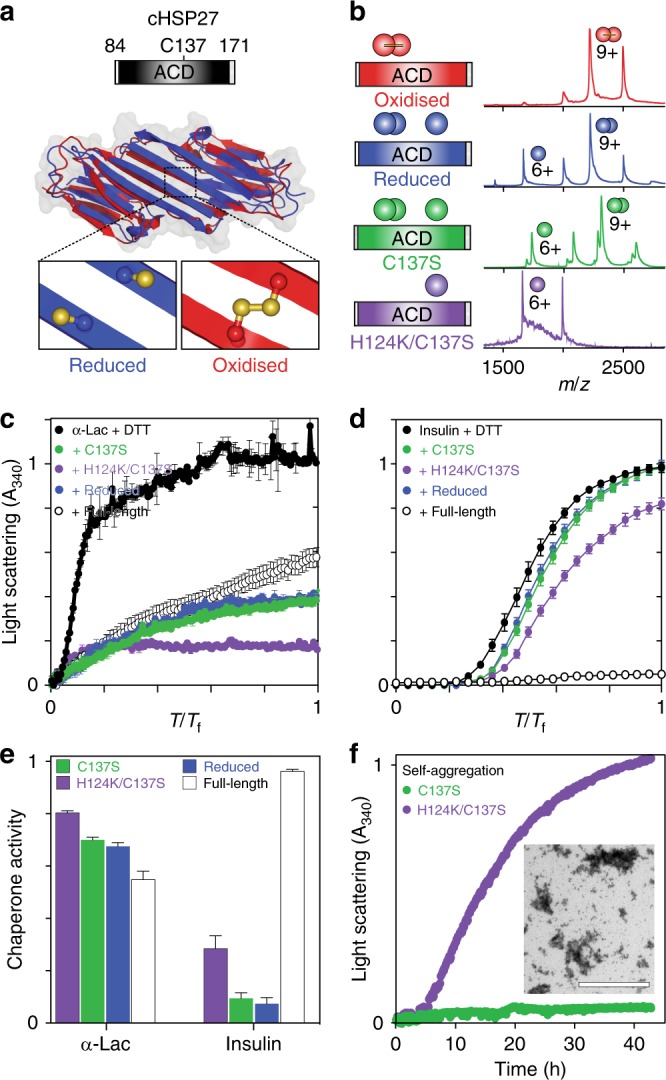
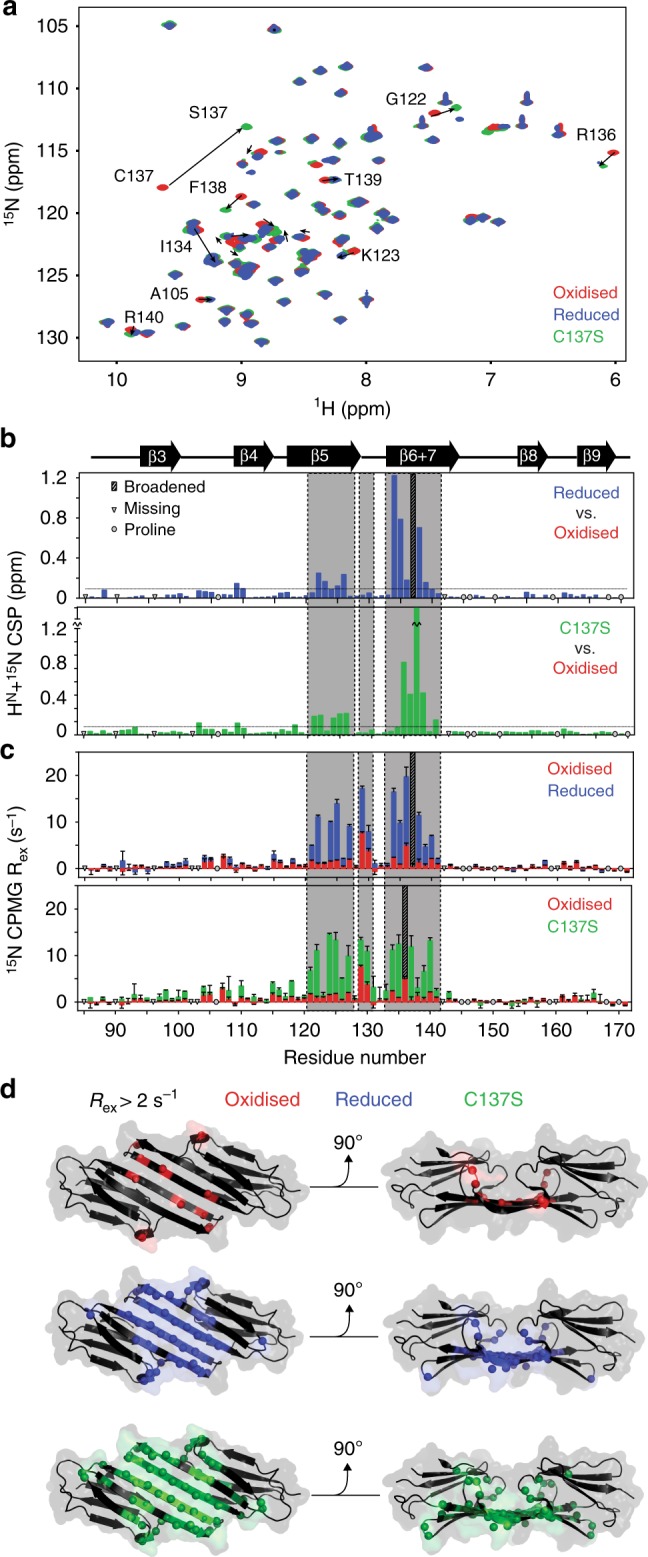
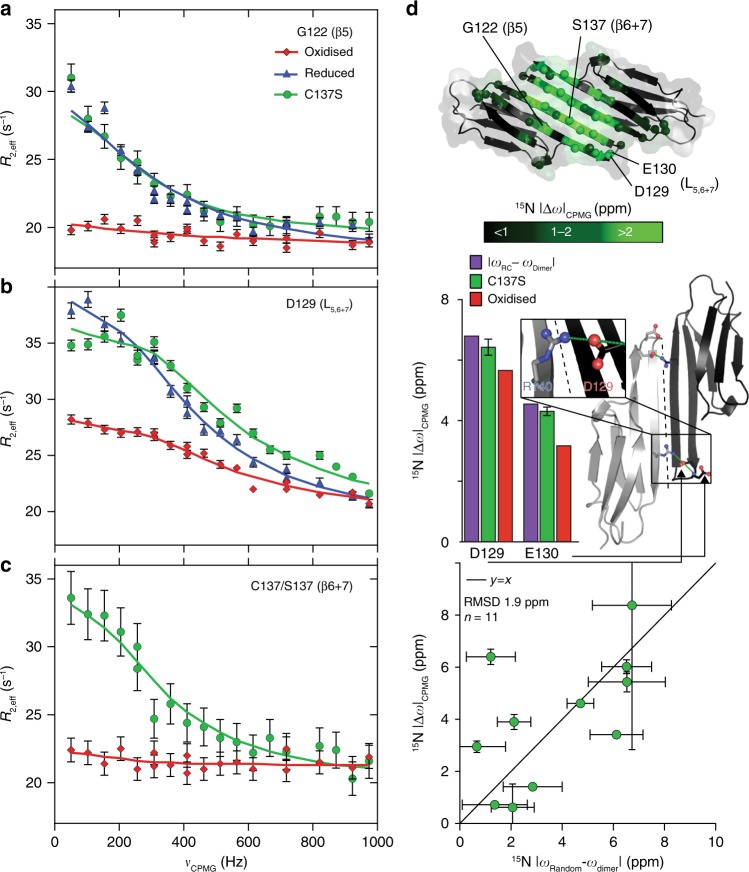
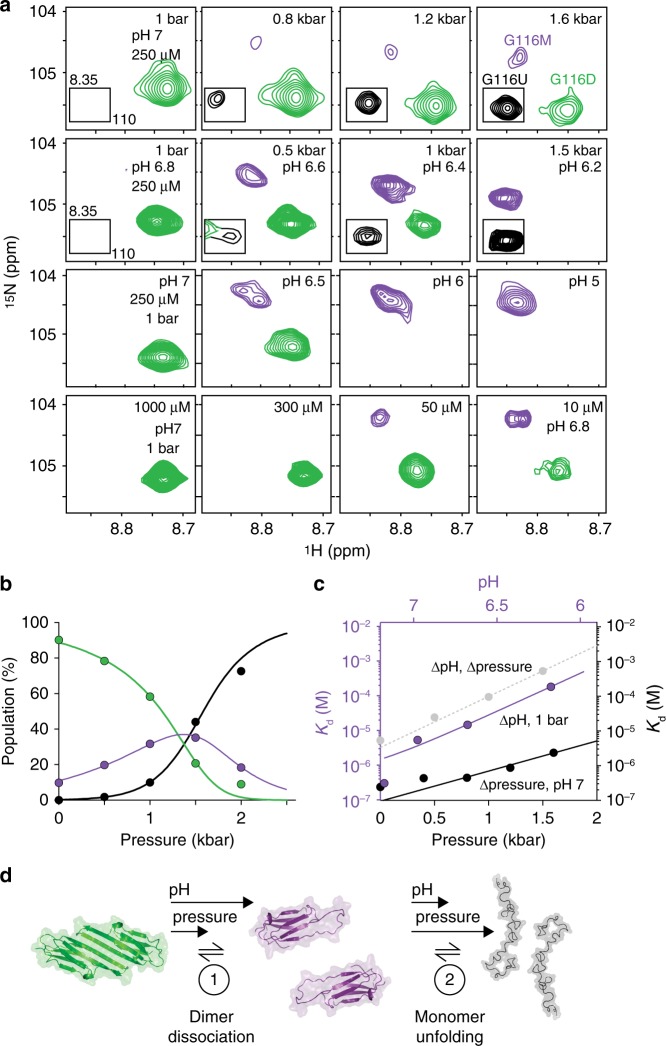
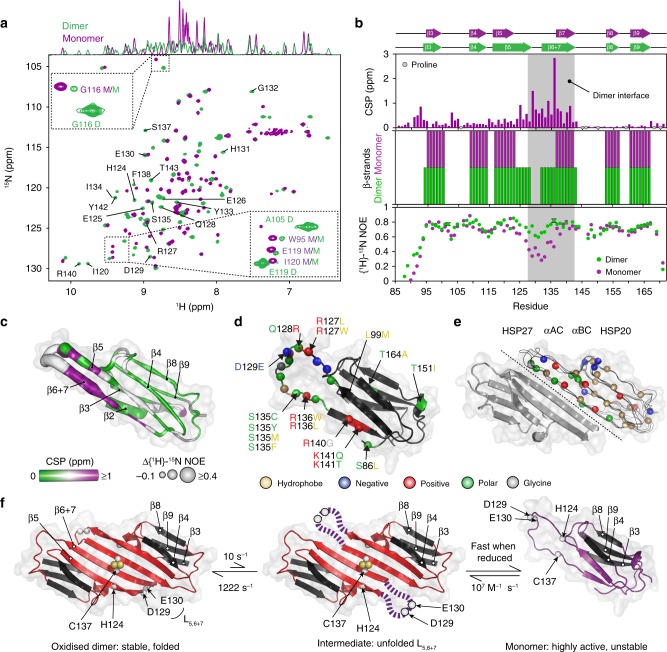
The small heat-shock protein HSP27 is a redox-sensitive molecular chaperone that is expressed throughout the human body. Here, we describe redox-induced changes to the structure, dynamics, and function of HSP27 and its conserved α-crystallin domain (ACD). While HSP27 assembles into oligomers, we show that the monomers formed upon reduction are highly active chaperones in vitro, but are susceptible to self-aggregation. By using relaxation dispersion and high-pressure nuclear magnetic resonance (NMR) spectroscopy, we observe that the pair of β-strands that mediate dimerisation partially unfold in the monomer. We note that numerous HSP27 mutations associated with inherited neuropathies cluster to this dynamic region. High levels of sequence conservation in ACDs from mammalian sHSPs suggest that the exposed, disordered interface present in free monomers or oligomeric subunits may be a general, functional feature of sHSPs.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Kampinga, H. H., de Boer, R. & Beerstra, N. in The Big Book on Small Heat Shock Proteins (Tanguay, R. M. & Hightower, L. E. eds.), 3–26 (Springer International Publishing, Cham, Switzerland, 2015). https://www.springer.com/gp/book/9783319160764
-
- Vos MJ, Kanon B, Kampinga HH. HSPB7 is a SC35 speckle resident small heat shock protein. Biochim. Biophys. Acta - Mol. Cell Res. 2009;1793:1343–1353. - PubMed
-
- Yu AL, et al. Oxidative stress and TGF-β2 increase heat shock protein 27 expression in human optic nerve head astrocytes. Investig. Opthalmology Vis. Sci. 2008;49:5403. - PubMed
-
- Weindruch R, Prolla TA, Lee CK. Gene-expression profile of the ageing brain in mice. Nat. Genet. 2000;25:294–297. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
