

L1cam-mediated developmental processes of the nervous system are differentially regulated by proteolytic processing
- PMID: 30842511
- PMCID: PMC6403279
- DOI: 10.1038/s41598-019-39884-x
L1cam-mediated developmental processes of the nervous system are differentially regulated by proteolytic processing
Abstract
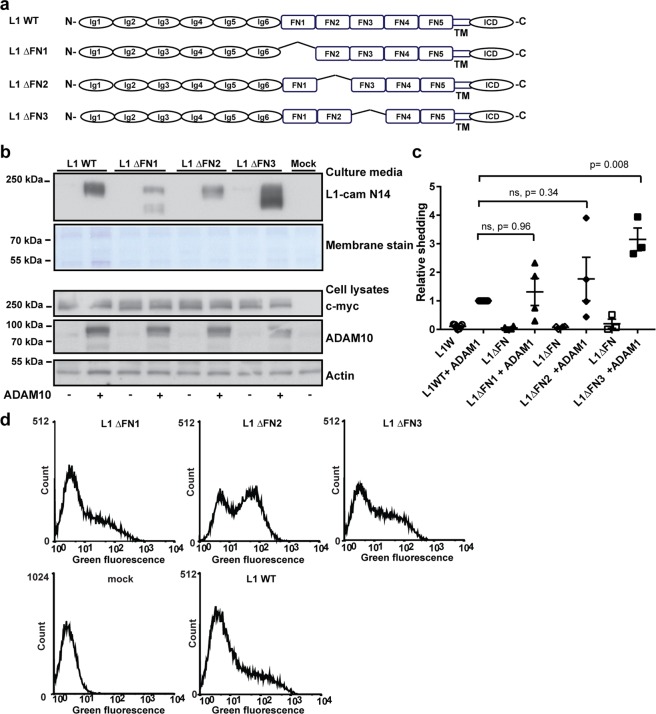
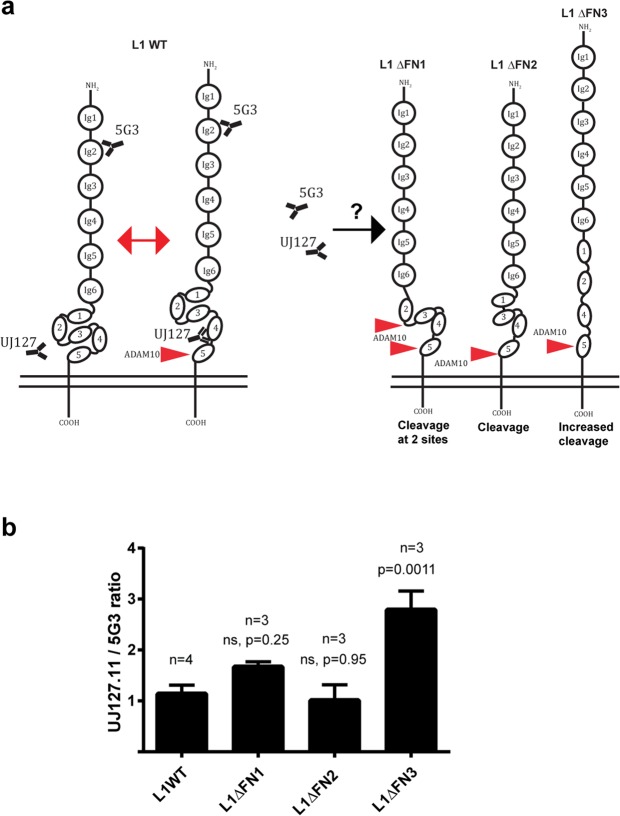
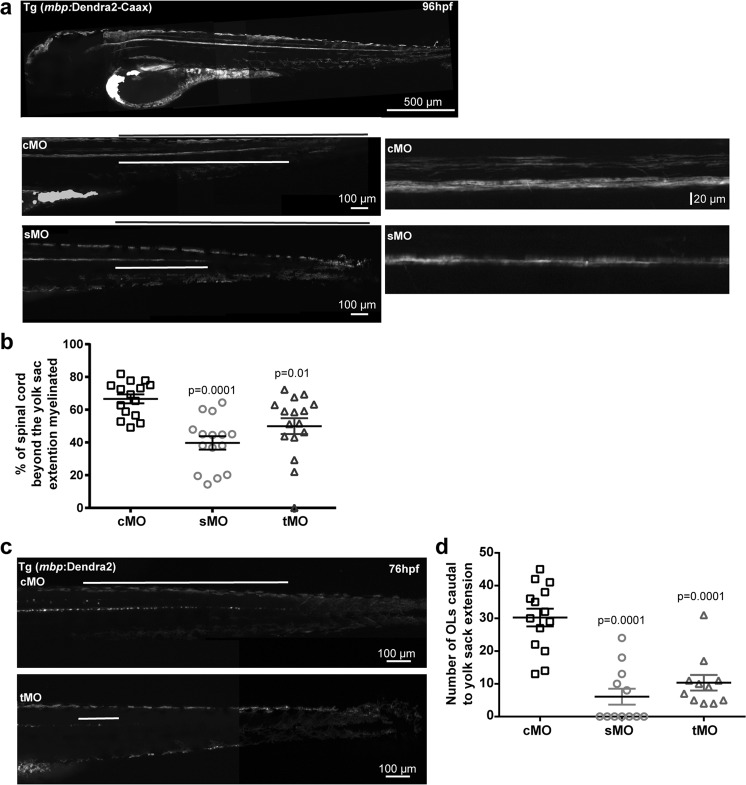
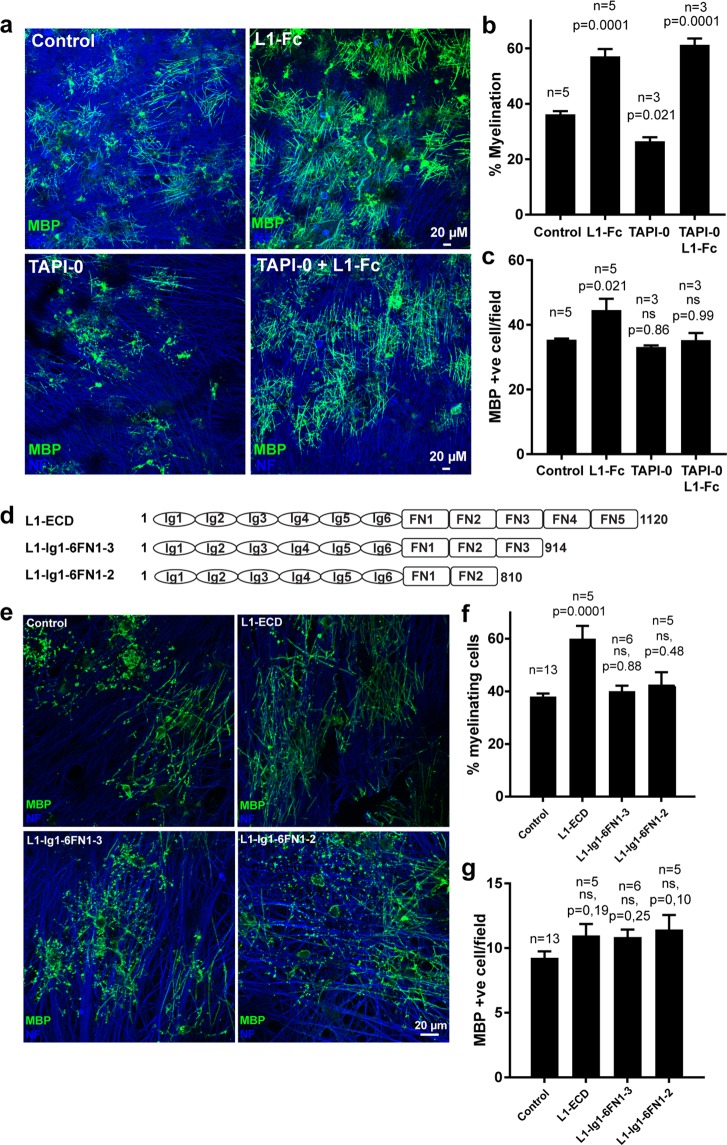
Normal brain development depends on tight temporal and spatial regulation of connections between cells. Mutations in L1cam, a member of the immunoglobulin (Ig) superfamily that mediate cell-cell contacts through homo- and heterophilic interactions, are associated with several developmental abnormalities of the nervous system, including mental retardation, limb spasticity, hydrocephalus, and corpus callosum aplasia. L1cam has been reported to be shed from the cell surface, but the significance of this during different phases of brain development is unknown. We here show that ADAM10-mediated shedding of L1cam is regulated by its fibronectin type III (FNIII) domains. Specifically, the third FNIII domain is important for maintaining a conformation where access to a membrane proximal cleavage site is restricted. To define the role of ADAM10/17/BACE1-mediated shedding of L1cam during brain development, we used a zebrafish model system. Knockdown of the zebrafish, l1camb, caused hydrocephalus, defects in axonal outgrowth, and myelination abnormalities. Rescue experiments with proteinase-resistant and soluble L1cam variants showed that proteolytic cleavage is not required for normal axonal outgrowth and development of the ventricular system. In contrast, metalloproteinase-mediated shedding is required for efficient myelination, and only specific fragments are able to mediate this stimulatory function of the shedded L1cam.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Stumpel, C. & Vos, Y. J. In GeneReVIEws((R)) (eds Adam, M. P. et al.) (1993).
-
- Demyanenko GP, Tsai AY, Maness PF. Abnormalities in Neuronal Process Extension, Hippocampal Development, and the Ventricular System of L1 Knockout Mice. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 1999;19:4907–4920. doi: 10.1523/JNEUROSCI.19-12-04907.1999. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
