

The IAP antagonist birinapant potentiates bortezomib anti-myeloma activity in vitro and in vivo
- PMID: 30845975
- PMCID: PMC6407248
- DOI: 10.1186/s13045-019-0713-x
The IAP antagonist birinapant potentiates bortezomib anti-myeloma activity in vitro and in vivo
Abstract
Background: Mechanisms by which Smac mimetics (SMs) interact with proteasome inhibitors (e.g., bortezomib) are largely unknown, particularly in multiple myeloma (MM), a disease in which bortezomib represents a mainstay of therapy.
Methods: Interactions between the clinically relevant IAP (inhibitor of apoptosis protein) antagonist birinapant (TL32711) and the proteasome inhibitor bortezomib were investigated in multiple myeloma (MM) cell lines and primary cells, as well as in vivo models. Induction of apoptosis and changes in gene and protein expression were monitored using MM cell lines and confirmed in primary MM cell populations. Genetically modified cells (e.g., exhibiting shRNA knockdown or ectopic expression) were employed to evaluate the functional significance of birinapant/bortezomib-induced changes in protein levels. A MM xenograft model was used to evaluate the in vivo activity of the birinapant/bortezomib regimen.
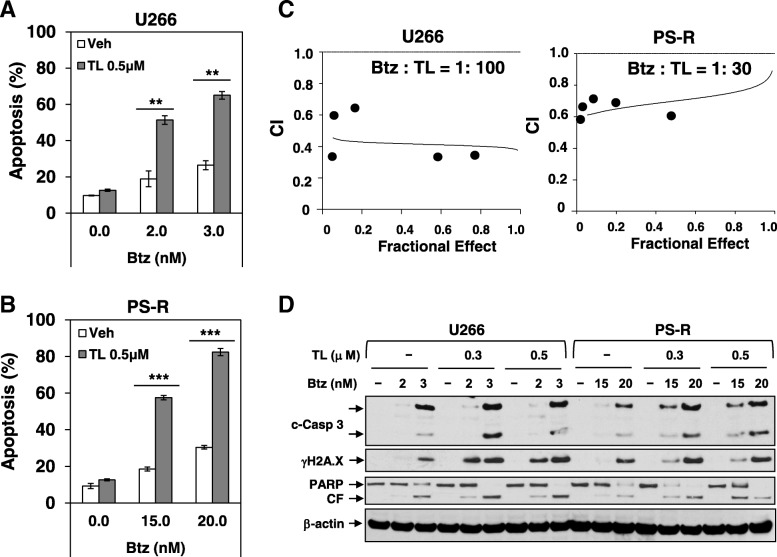
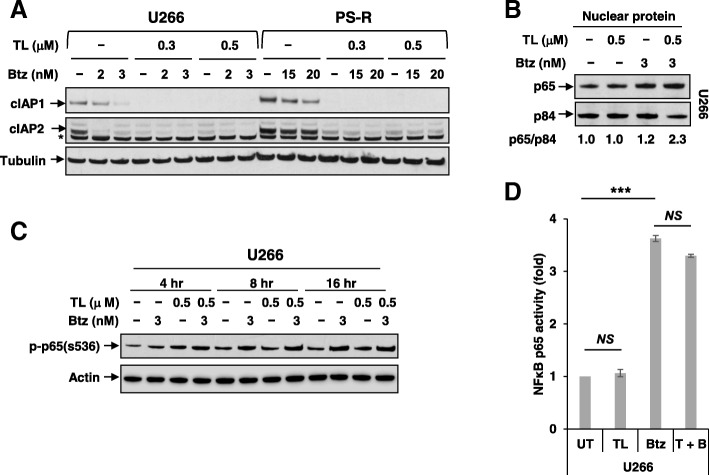
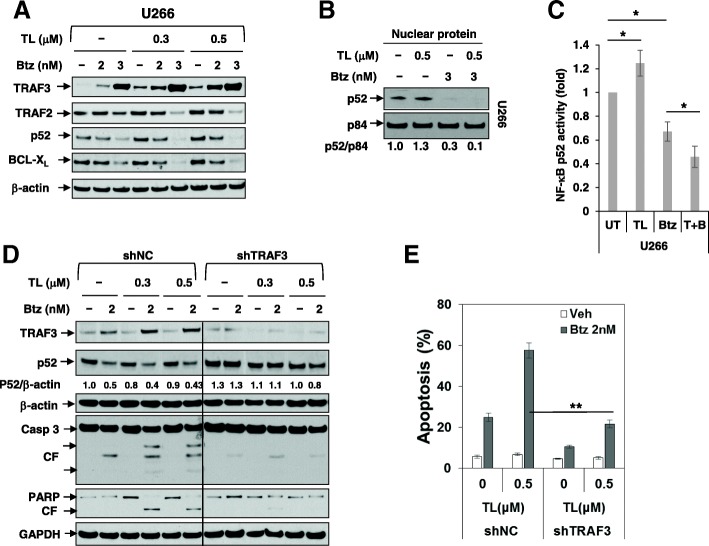
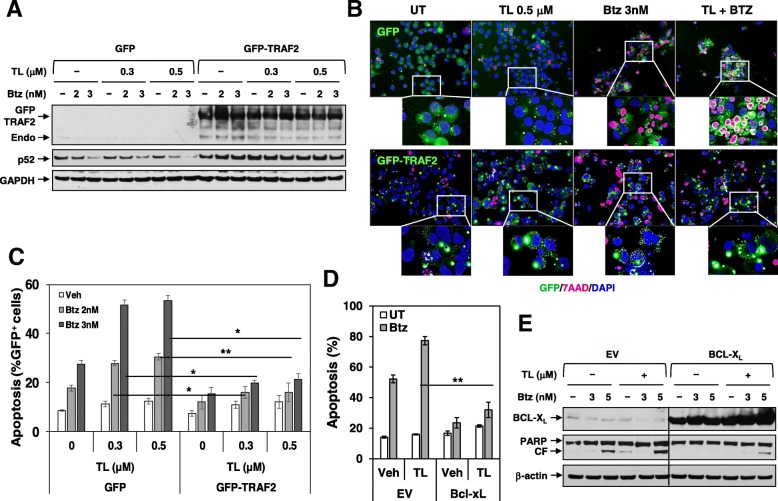
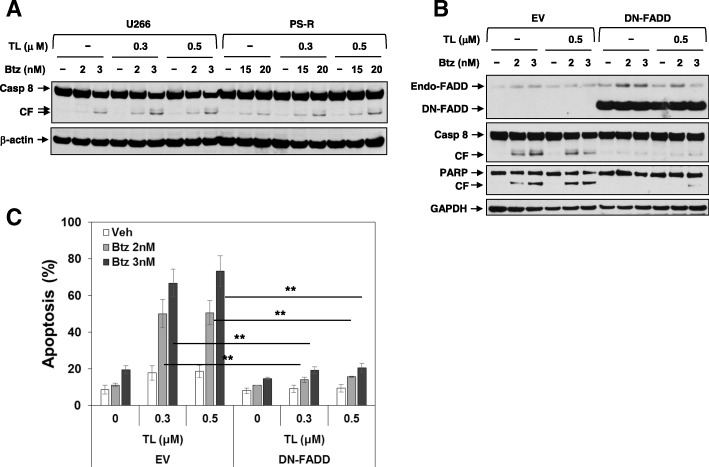
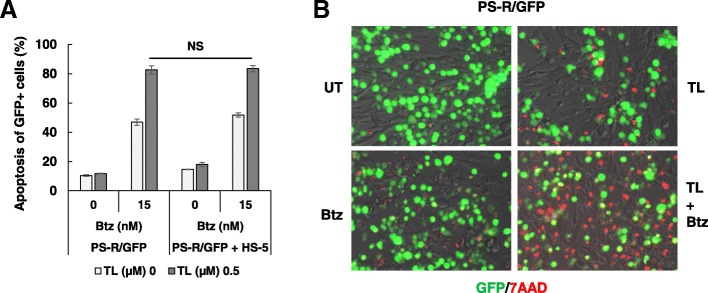
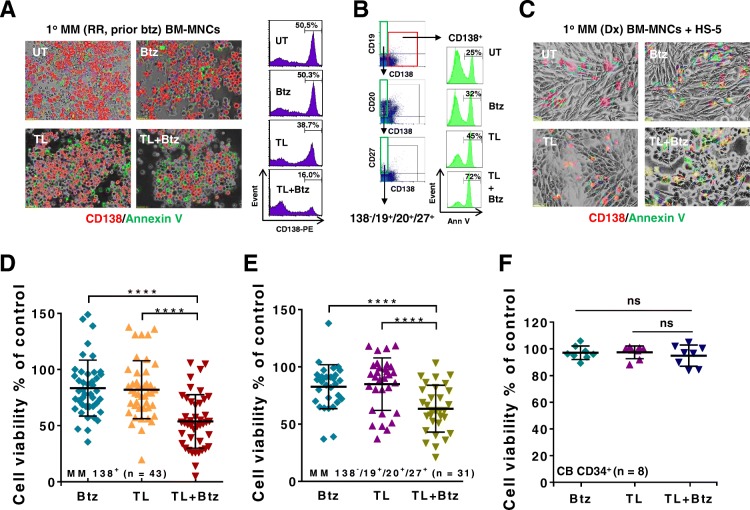
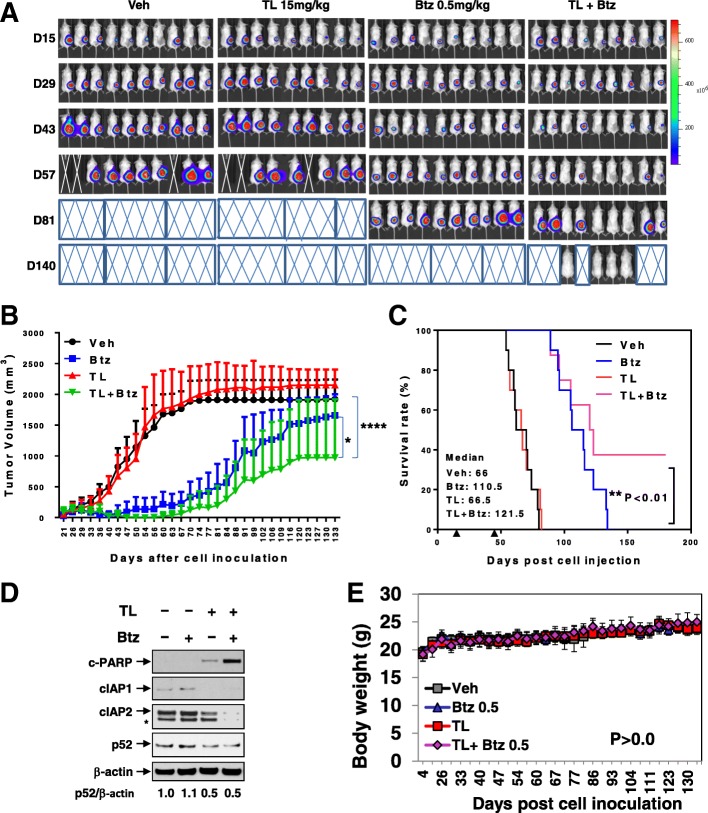
Results: Birinapant and bortezomib synergistically induced apoptosis in diverse cell lines, including bortezomib-resistant cells (PS-R). The regimen robustly downregulated cIAP1/2 but not the canonical NF-κB pathway, reflected by p65 phosphorylation and nuclear accumulation. In contrast, the bortezomib/birinapant regimen upregulated TRAF3, downregulated TRAF2, and diminished p52 processing and BCL-XL expression, consistent with disruption of the non-canonical NF-κB pathway. TRAF3 knockdown, ectopic TRAF2, or BCL-XL expression significantly diminished birinapant/bortezomib toxicity. The regimen sharply increased extrinsic apoptotic pathway activation, and cells expressing dominant-negative FADD or caspase-8 displayed markedly reduced birinapant/bortezomib sensitivity. Primary CD138+ (n = 43) and primitive MM populations (CD138-/19+/20+/27+; n = 31) but not normal CD34+ cells exhibited significantly enhanced toxicity with combined treatment (P < 0.0001). The regimen was also fully active in the presence of HS-5 stromal cells or growth factors (e.g., IL-6 and VEGF). Finally, the regimen was well tolerated and significantly increased survival (P < 0.05 and P < 0.001) compared to single agents in a MM xenograft model. Combined treatment also downregulated cIAP1/2 and p52 while increasing PARP cleavage in MM cells in vivo.
Conclusions: Our data suggest that birinapant and bortezomib interact synergistically in MM cells, including those resistant to bortezomib, through inactivation of the non-canonical NF-κB and activation of the extrinsic apoptotic pathway both in vitro and in vivo. They also argue that a strategy combining cIAP antagonists and proteasome inhibitors warrants attention in MM.
Keywords: Bortezomib; IAP antagonist; Multiple myeloma; NF-κB.
Conflict of interest statement
Ethics approval and consent to participate
Mice were bred, treated, and maintained under pathogen-free conditions in-house under Virginia Commonwealth University IACUC-approved protocols and as mandated by federal law and regulations. The experimental protocol was conducted in accordance with the recommendations of the Guide for Care and Use of Laboratory Animals with respect to restraint, husbandry, surgical procedures, feed and fluid regulation, and veterinary care. The animal care and use program at Virginia Commonwealth University is accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
Mechanisms underlying synergism between circularized tumor necrosis factor-related apoptosis inducing ligand and bortezomib in bortezomib-sensitive or -resistant myeloma cells.Hematol Oncol. 2022 Dec;40(5):999-1008. doi: 10.1002/hon.3045. Epub 2022 Jul 14. Hematol Oncol. 2022. PMID: 35789025 Free PMC article.
-
IAP and HDAC inhibitors interact synergistically in myeloma cells through noncanonical NF-κB- and caspase-8-dependent mechanisms.Blood Adv. 2021 Oct 12;5(19):3776-3788. doi: 10.1182/bloodadvances.2020003597. Blood Adv. 2021. PMID: 34464977 Free PMC article.
-
Birinapant (TL32711), a bivalent SMAC mimetic, targets TRAF2-associated cIAPs, abrogates TNF-induced NF-κB activation, and is active in patient-derived xenograft models.Mol Cancer Ther. 2014 Apr;13(4):867-79. doi: 10.1158/1535-7163.MCT-13-0798. Epub 2014 Feb 21. Mol Cancer Ther. 2014. PMID: 24563541
-
The role of nuclear factor-kappaB in the biology and treatment of multiple myeloma.Semin Oncol. 2001 Dec;28(6):626-33. doi: 10.1016/s0093-7754(01)90036-3. Semin Oncol. 2001. PMID: 11740821 Review.
-
The IAP Protein Family, SMAC Mimetics and Cancer Treatment.Crit Rev Oncog. 2016;21(3-4):185-202. doi: 10.1615/CritRevOncog.2016017032. Crit Rev Oncog. 2016. PMID: 27915971 Review.
Cited by
-
Protein-Functionalized Microgel for Multiple Myeloma Cells' 3D Culture.Biomedicines. 2022 Nov 3;10(11):2797. doi: 10.3390/biomedicines10112797. Biomedicines. 2022. PMID: 36359316 Free PMC article.
-
Dual-Targeted Therapy Circumvents Non-Genetic Drug Resistance to Targeted Therapy.Front Oncol. 2022 Apr 27;12:859455. doi: 10.3389/fonc.2022.859455. eCollection 2022. Front Oncol. 2022. PMID: 35574302 Free PMC article. Review.
-
Regulatory role of E3 ubiquitin ligases in multiple myeloma: from molecular mechanisms to therapeutic strategies.Front Cell Dev Biol. 2025 Jul 30;13:1620097. doi: 10.3389/fcell.2025.1620097. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40809696 Free PMC article. Review.
-
Alternative NF-κB Signaling Discriminates Induction of the Tumor Marker Fascin by the Viral Oncoproteins Tax-1 and Tax-2 of Human T-Cell Leukemia Viruses.Cancers (Basel). 2022 Jan 21;14(3):537. doi: 10.3390/cancers14030537. Cancers (Basel). 2022. PMID: 35158803 Free PMC article.
-
Mechanisms underlying synergism between circularized tumor necrosis factor-related apoptosis inducing ligand and bortezomib in bortezomib-sensitive or -resistant myeloma cells.Hematol Oncol. 2022 Dec;40(5):999-1008. doi: 10.1002/hon.3045. Epub 2022 Jul 14. Hematol Oncol. 2022. PMID: 35789025 Free PMC article.
References
-
- Deckert J, Wetzel MC, Bartle LM, Skaletskaya A, Goldmacher VS, Vallee F, et al. SAR650984, a novel humanized CD38-targeting antibody, demonstrates potent antitumor activity in models of multiple myeloma and other CD38+ hematologic malignancies. Clin Cancer Res. 2014;20:4574–4583. doi: 10.1158/1078-0432.CCR-14-0695. - DOI - PubMed
-
- Raab M, Goldschmidt H, Agis H, Blau I, Einsele H, Engelhardt M, et al. A phase I/Iia study of the human anti-Cd38 antibody Mor202 (Mor03087) in relapsed or refractory multiple myeloma. Haematologica. 2015;100:312–313.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
