

Commensal Staphylococci Influence Staphylococcus aureus Skin Colonization and Disease
- PMID: 30846311
- PMCID: PMC7176043
- DOI: 10.1016/j.tim.2019.01.008
Commensal Staphylococci Influence Staphylococcus aureus Skin Colonization and Disease
Abstract
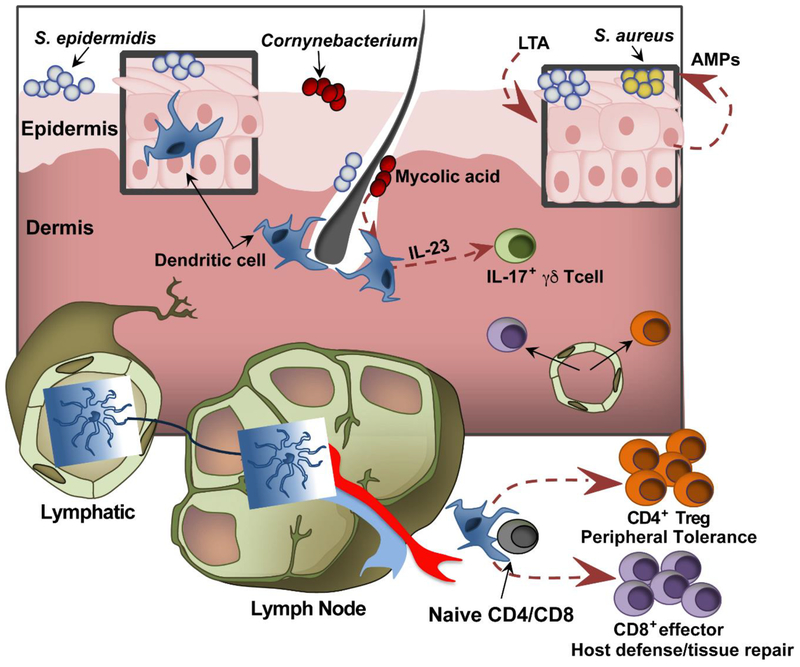
Commensal organisms that constitute the skin microbiota play a pivotal role in the orchestration of cutaneous homeostasis and immune competence. This balance can be promptly offset by the expansion of the opportunistic pathogen Staphylococcus aureus, which is responsible for the majority of bacterial skin infections. S. aureus carriage is also known to be a precondition for its transmission and pathogenesis. Recent reports suggest that skin-dwelling coagulase-negative staphylococci (CoNS) can prime the skin immune system to limit the colonization potential of invaders, and they can directly compete through production of antimicrobial molecules or through signaling antagonism. We review recent advances in these CoNS colonization resistance mechanisms, which may serve to aid development of pharmacologic and probiotic intervention strategies to limit S. aureus skin colonization and disease.
Keywords: MRSA; Staphylococcus aureus; colonization; host protection; quorum sensing; skin microbiota.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures



References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
