

Separate Pathways Contribute to the Herbivore-Induced Formation of 2-Phenylethanol in Poplar
- PMID: 30846485
- PMCID: PMC6548255
- DOI: 10.1104/pp.19.00059
Separate Pathways Contribute to the Herbivore-Induced Formation of 2-Phenylethanol in Poplar
Abstract
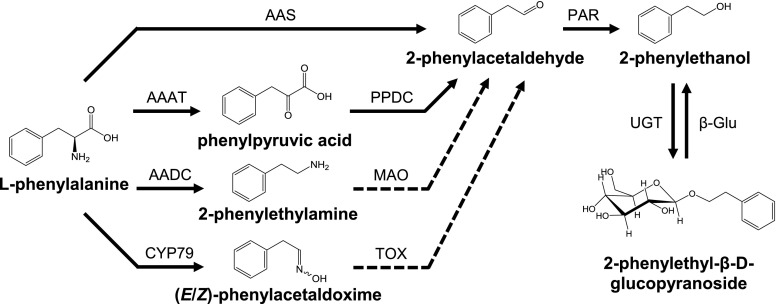
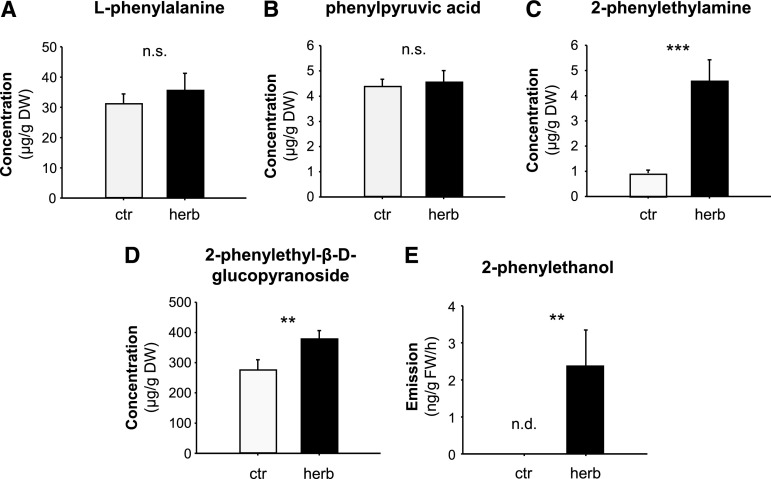
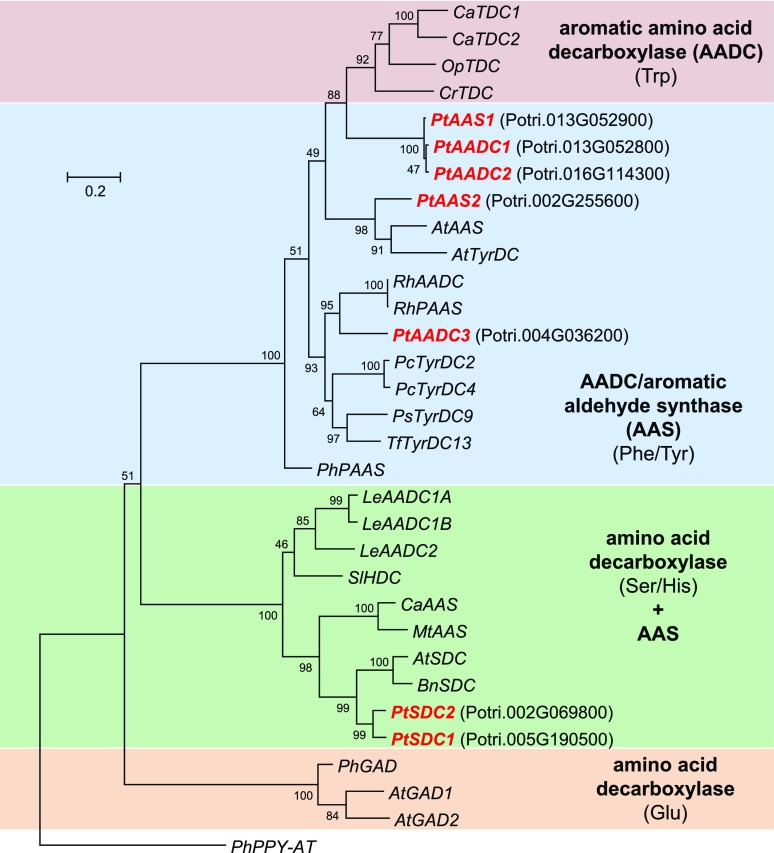
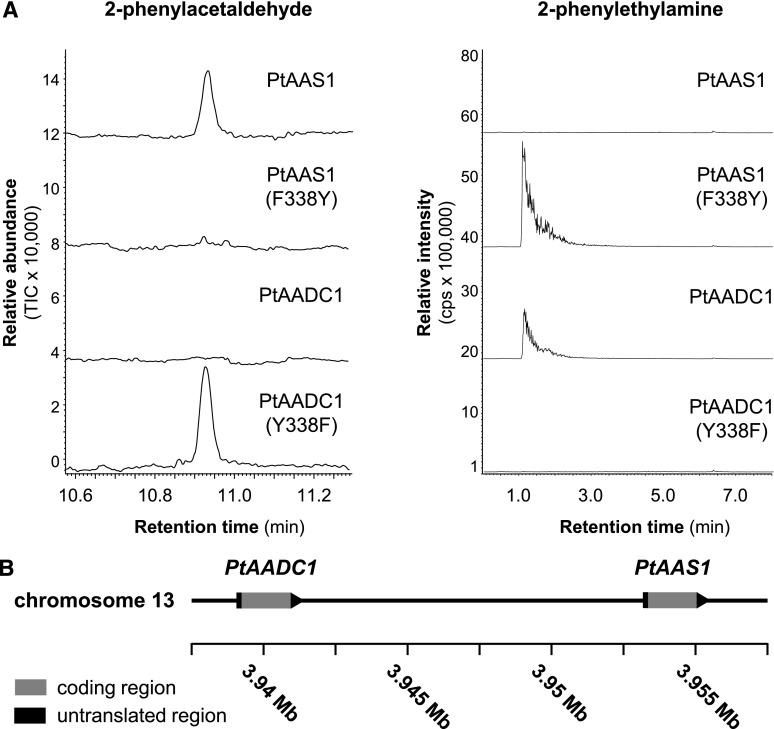
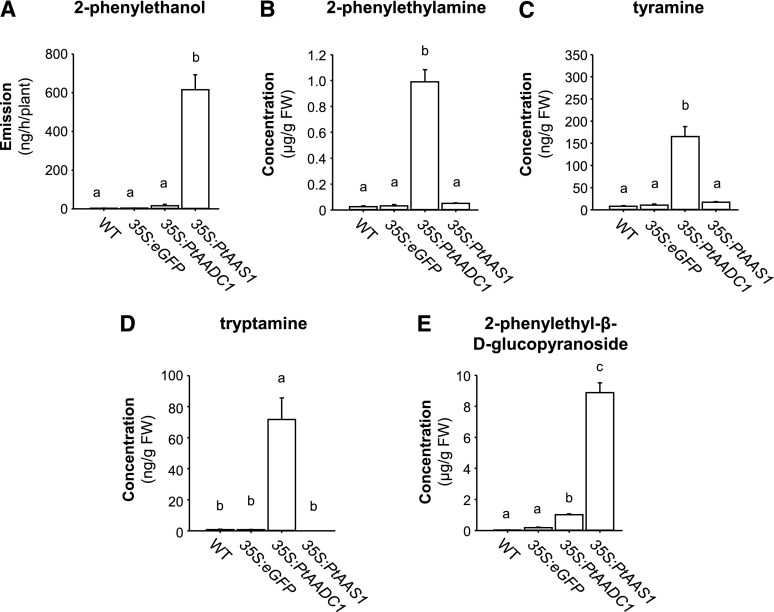
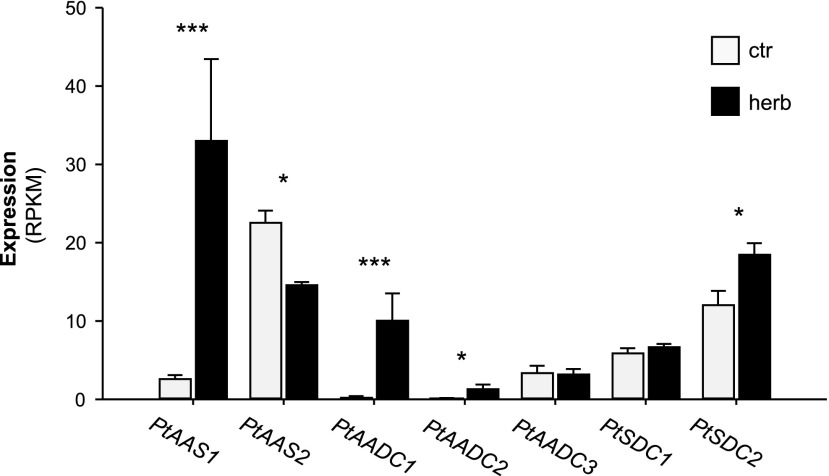
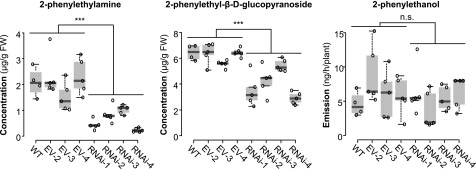
Upon herbivory, the tree species western balsam poplar (Populus trichocarpa) produces a variety of Phe-derived metabolites, including 2-phenylethylamine, 2-phenylethanol, and 2-phenylethyl-β-d-glucopyranoside. To investigate the formation of these potential defense compounds, we functionally characterized aromatic l-amino acid decarboxylases (AADCs) and aromatic aldehyde synthases (AASs), which play important roles in the biosynthesis of specialized aromatic metabolites in other plants. Heterologous expression in Escherichia coli and Nicotiana benthamiana showed that all five AADC/AAS genes identified in the P trichocarpa genome encode active enzymes. However, only two genes, PtAADC1 and PtAAS1, were significantly upregulated after leaf herbivory. Despite a sequence similarity of ∼96%, PtAADC1 and PtAAS1 showed different enzymatic functions and converted Phe into 2-phenylethylamine and 2-phenylacetaldehyde, respectively. The activities of both enzymes were interconvertible by switching a single amino acid residue in their active sites. A survey of putative AADC/AAS gene pairs in the genomes of other plants suggests an independent evolution of this function-determining residue in different plant families. RNA interference -mediated-downregulation of AADC1 in gray poplar (Populus × canescens) resulted in decreased accumulation of 2-phenylethylamine and 2-phenylethyl-β-d-glucopyranoside, whereas the emission of 2-phenylethanol was not influenced. To investigate the last step of 2-phenylethanol formation, we identified and characterized two P trichocarpa short-chain dehydrogenases, PtPAR1 and PtPAR2, which were able to reduce 2-phenylacetaldehyde to 2-phenylethanol in vitro. In summary, 2-phenylethanol and its glucoside may be formed in multiple ways in poplar. Our data indicate that PtAADC1 controls the herbivore-induced formation of 2-phenylethylamine and 2-phenylethyl-β-d-glucopyranoside in planta, whereas PtAAS1 likely contributes to the herbivore-induced emission of 2-phenylethanol.
© 2019 American Society of Plant Biologists. All Rights Reserved.
Figures
Comment in
-
Do All Roads Lead to 2-Phenylethanol in Populus?Plant Physiol. 2019 Jun;180(2):693-694. doi: 10.1104/pp.19.00424. Plant Physiol. 2019. PMID: 31160522 Free PMC article. No abstract available.
References
-
- Arimura G, Huber DPW, Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa × deltoides): cDNA cloning, functional characterization, and patterns of gene expression of (−)-germacrene D synthase, PtdTPS1. Plant J 37: 603–616 - PubMed
-
- Behnke K, Ehlting B, Teuber M, Bauerfeind M, Louis S, Hänsch R, Polle A, Bohlmann J, Schnitzler JP (2007) Transgenic, non-isoprene emitting poplars don’t like it hot. Plant J 51: 485–499 - PubMed
-
- Bertoldi M, Gonsalvi M, Contestabile R, Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-dependent oxidative deaminase. J Biol Chem 277: 36357–36362 - PubMed
-
- Bieri S, Brachet A, Veuthey JL, Christen P (2006) Cocaine distribution in wild Erythroxylum species. J Ethnopharmacol 103: 439–447 - PubMed
-
- Campos L, Lisón P, López-Gresa MP, Rodrigo I, Zacarés L, Conejero V, Bellés JM (2014) Transgenic tomato plants overexpressing tyramine n-hydroxycinnamoyltransferase exhibit elevated hydroxycinnamic acid amide levels and enhanced resistance to Pseudomonas syringae. Mol Plant Microbe Interact 27: 1159–1169 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
