

Endocytosis of commensal antigens by intestinal epithelial cells regulates mucosal T cell homeostasis
- PMID: 30846568
- PMCID: PMC6708280
- DOI: 10.1126/science.aat4042
Endocytosis of commensal antigens by intestinal epithelial cells regulates mucosal T cell homeostasis
Abstract
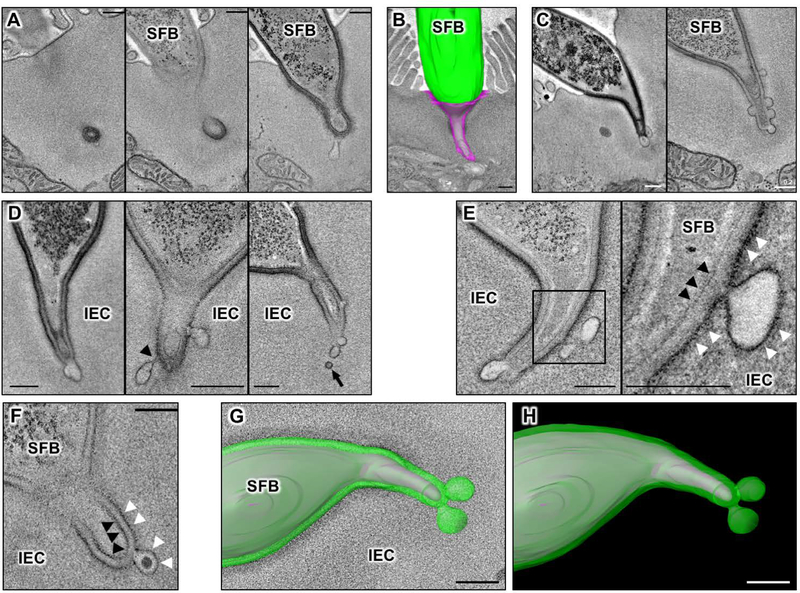
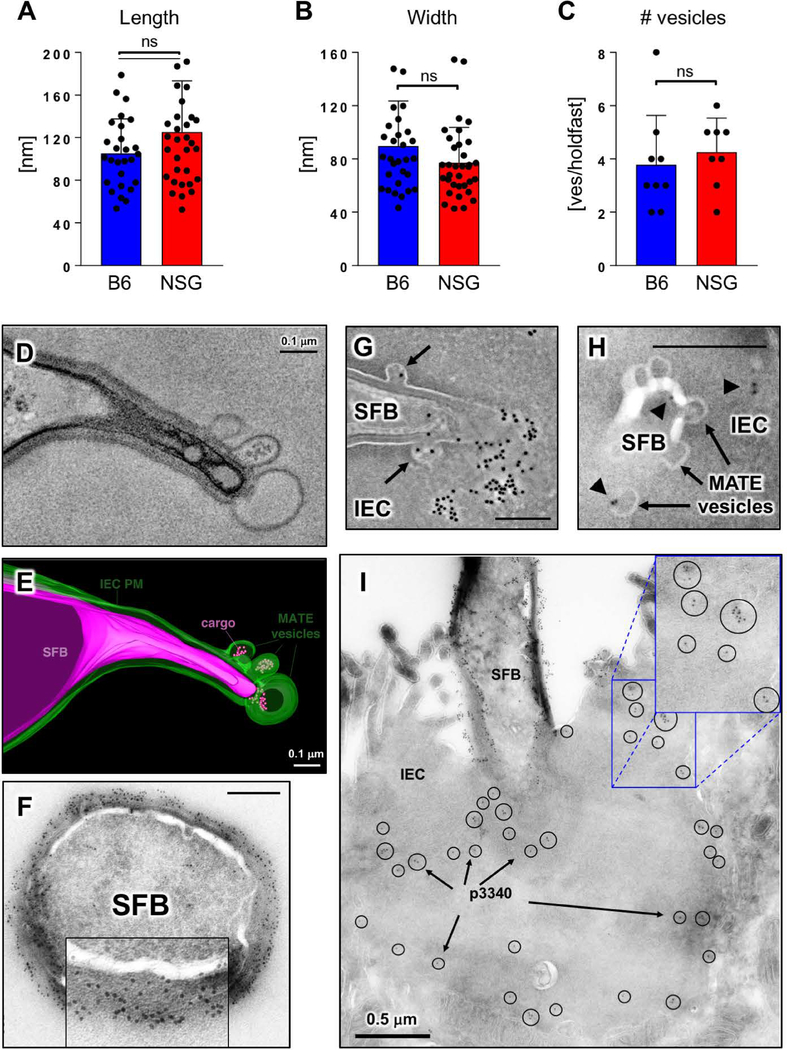
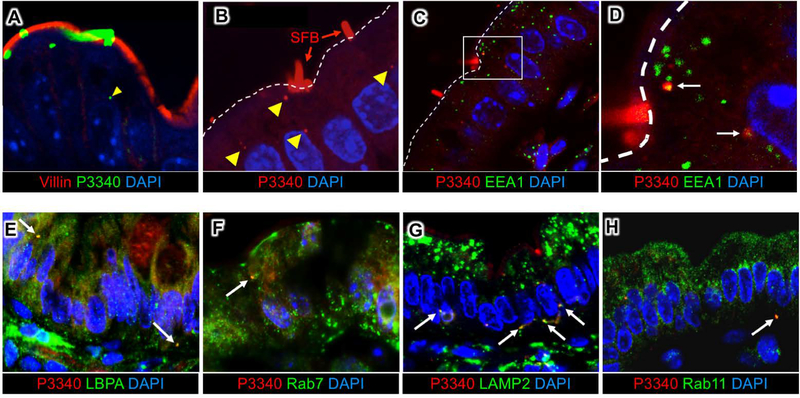
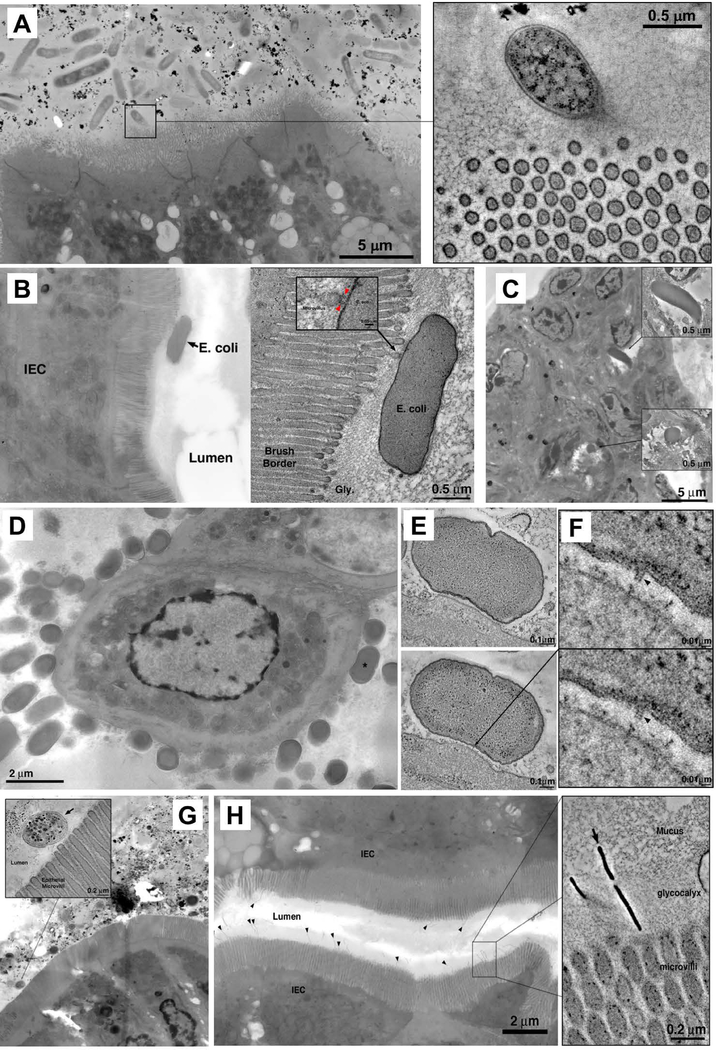
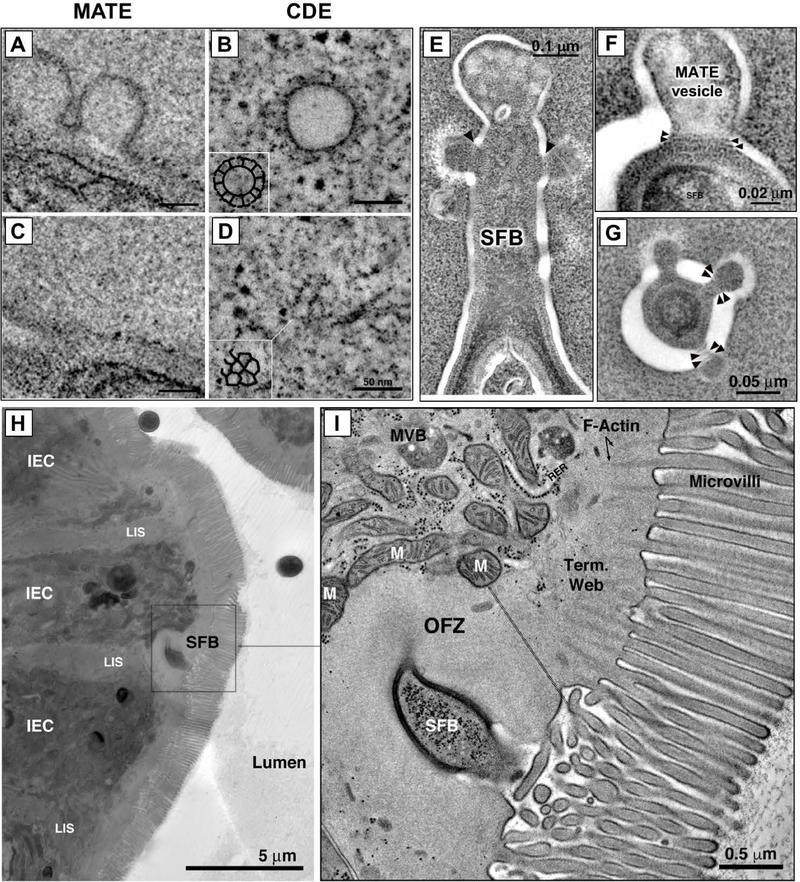
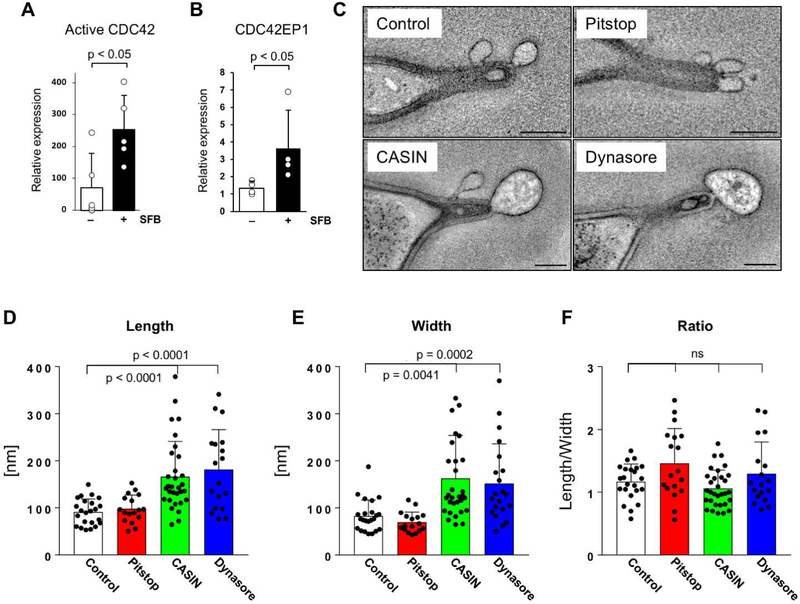
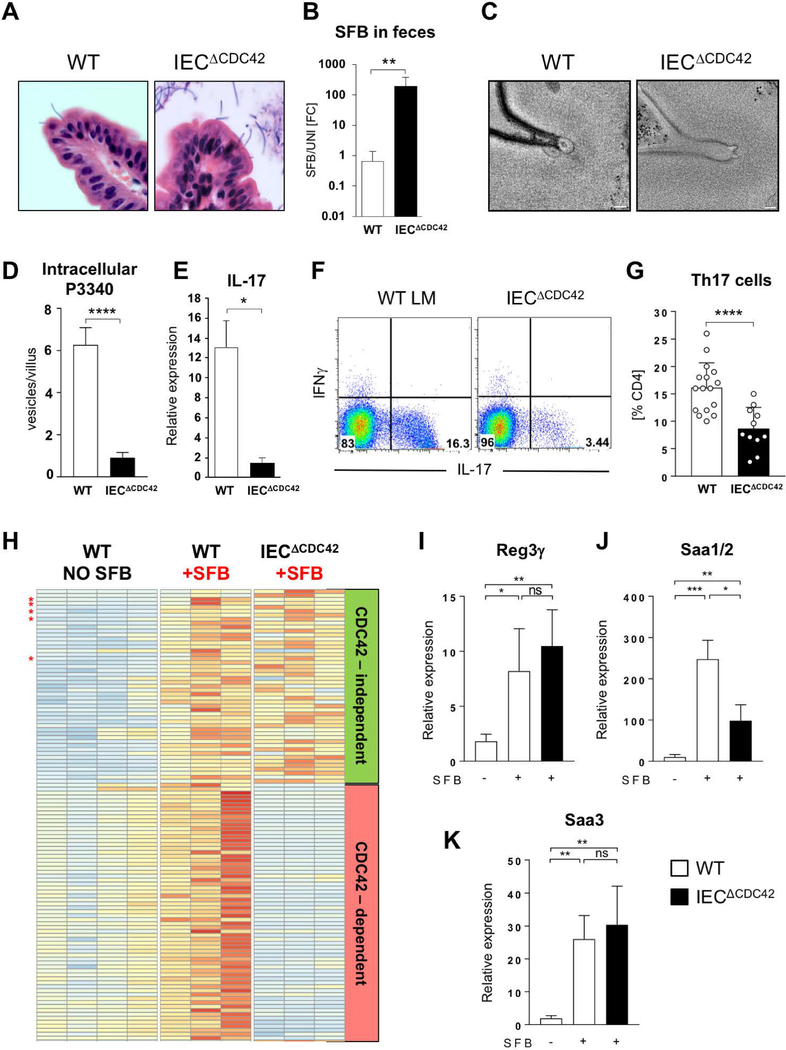
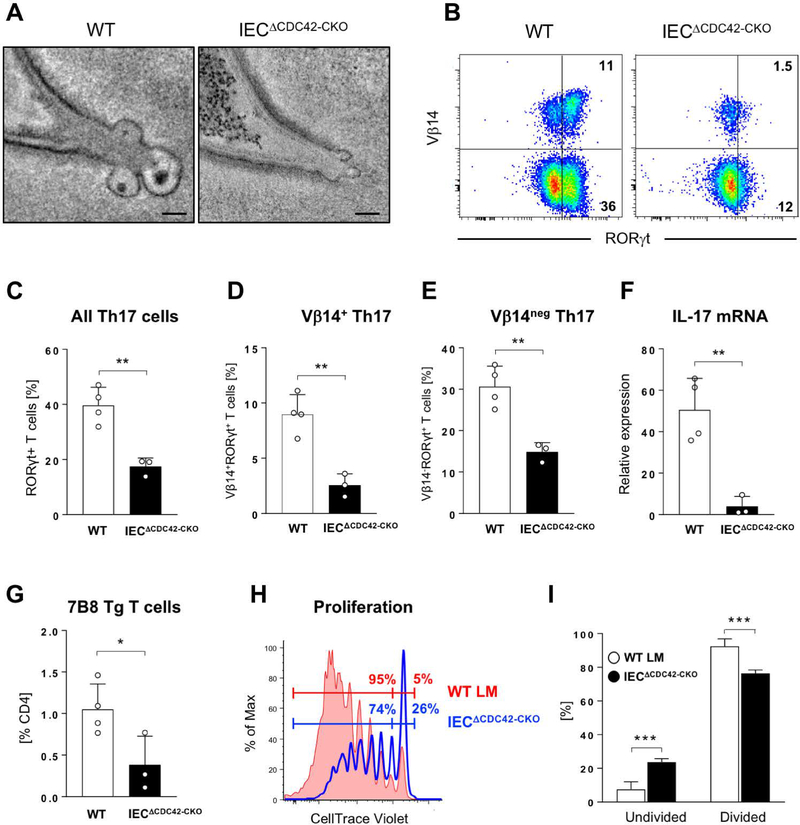
Commensal bacteria influence host physiology, without invading host tissues. We show that proteins from segmented filamentous bacteria (SFB) are transferred into intestinal epithelial cells (IECs) through adhesion-directed endocytosis that is distinct from the clathrin-dependent endocytosis of invasive pathogens. This process transfers microbial cell wall-associated proteins, including an antigen that stimulates mucosal T helper 17 (TH17) cell differentiation, into the cytosol of IECs in a cell division control protein 42 homolog (CDC42)-dependent manner. Removal of CDC42 activity in vivo led to disruption of endocytosis induced by SFB and decreased epithelial antigen acquisition, with consequent loss of mucosal TH17 cells. Our findings demonstrate direct communication between a resident gut microbe and the host and show that under physiological conditions, IECs acquire antigens from commensal bacteria for generation of T cell responses to the resident microbiota.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
