

NtMYB4 and NtCHS1 Are Critical Factors in the Regulation of Flavonoid Biosynthesis and Are Involved in Salinity Responsiveness
- PMID: 30846995
- PMCID: PMC6393349
- DOI: 10.3389/fpls.2019.00178
NtMYB4 and NtCHS1 Are Critical Factors in the Regulation of Flavonoid Biosynthesis and Are Involved in Salinity Responsiveness
Abstract
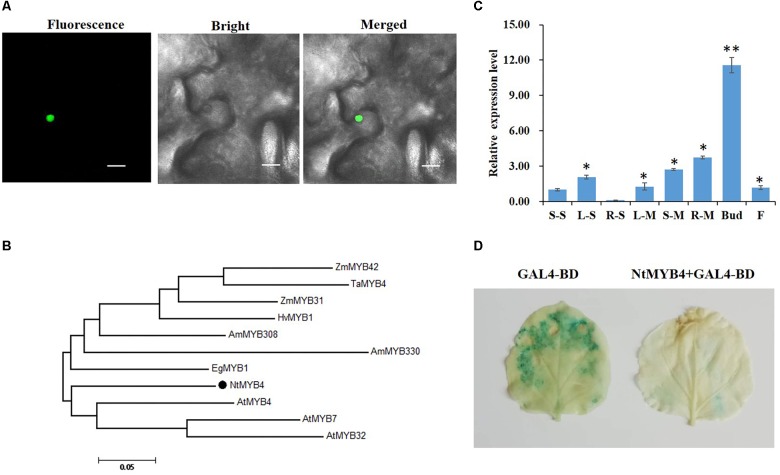
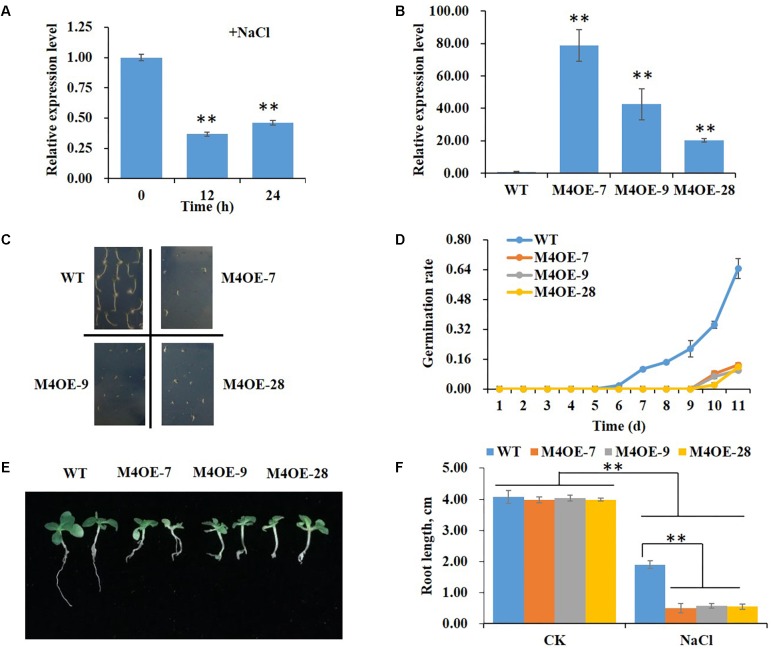
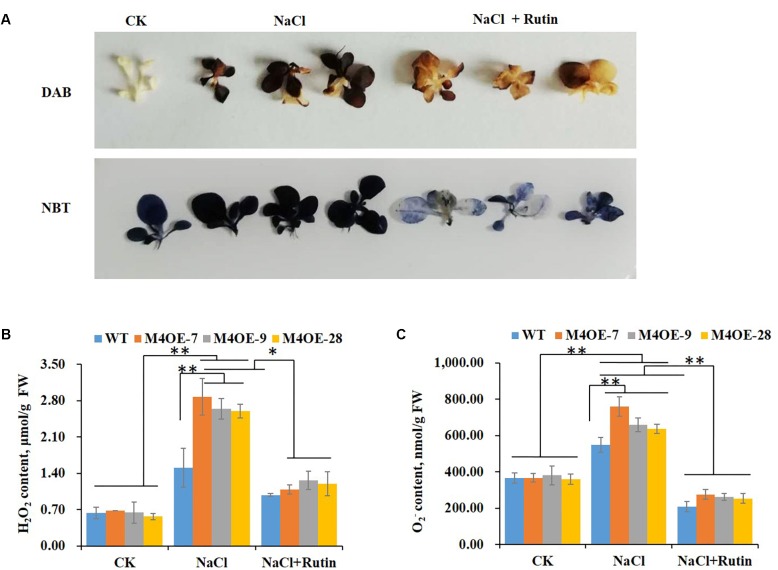
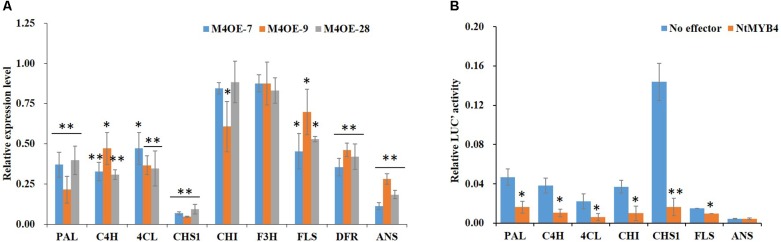
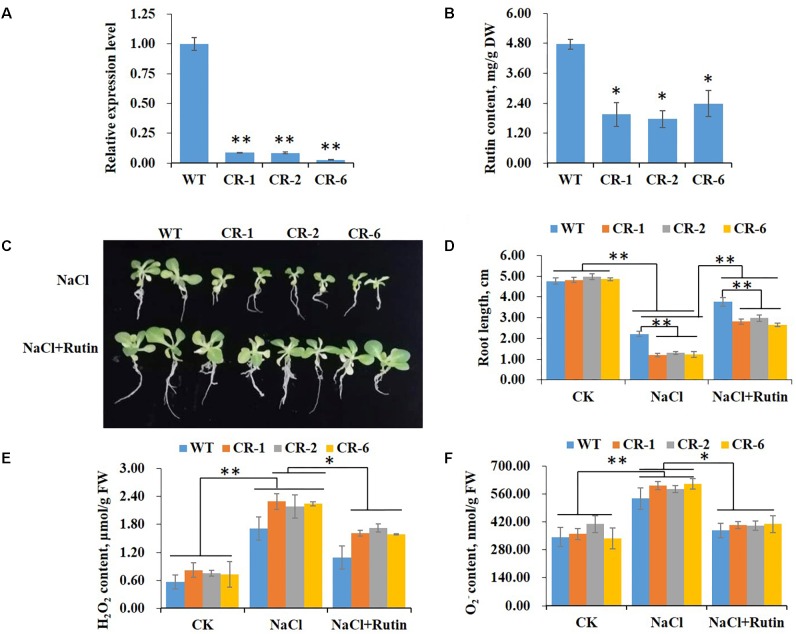
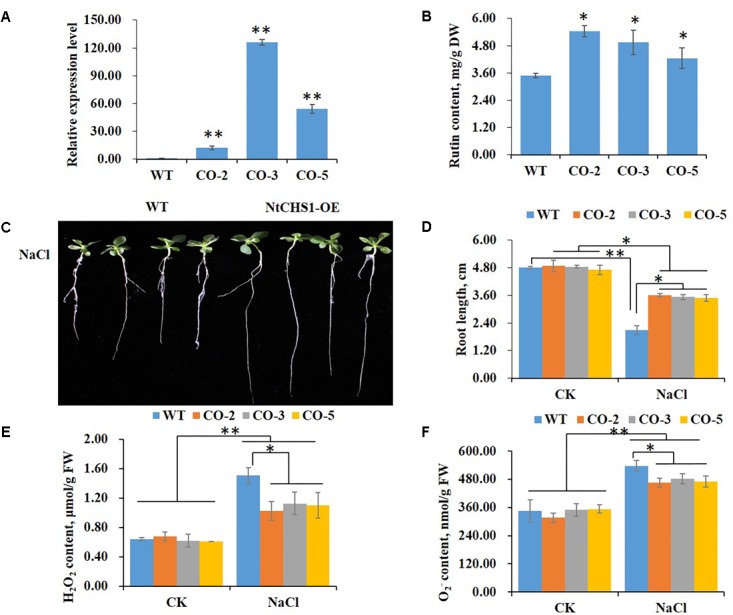
High levels of salinity induce serious oxidative damage in plants. Flavonoids, as antioxidants, have important roles in reactive oxygen species (ROS) scavenging. In the present study, the tobacco R2R3 MYB type repressor, NtMYB4, was isolated and characterized. The expression of NtMYB4 was suppressed by salinity. Overexpression of NtMYB4 reduced the salt tolerance in transgenic tobacco plants. NtMYB4 repressed the promoter activity of NtCHS1 and negatively regulated its expression. Rutin accumulation was significantly decreased in NtMYB4 overexpressing transgenic plants and NtCHS1 RNAi silenced transgenic plants. Moreover, high H2O2 and contents were detected in both types of rutin-reduced transgenic plants under high salt stress. In addition, exogenous rutin supplementation effectively scavenged ROS (H2O2 and ) and improved the salt tolerance of the rutin-reduced transgenic plants. In contrast, NtCHS1 overexpressing plants had increased rutin accumulation, lower H2O2 and contents, and higher tolerance to salinity. These results suggested that tobacco NtMYB4 acts as a salinity response repressor and negatively regulates NtCHS1 expression, which results in the reduced flavonoid accumulation and weakened ROS-scavenging ability under salt stress.
Keywords: NtCHS1; NtMYB4; ROS level; flavonoid pathway; salt stress.
Figures

Similar articles
-
GhFB15 is an F-box protein that modulates the response to salinity by regulating flavonoid biosynthesis.Plant Sci. 2024 Jan;338:111899. doi: 10.1016/j.plantsci.2023.111899. Epub 2023 Oct 20. Plant Sci. 2024. PMID: 37865208
-
The cotton WRKY transcription factor GhWRKY17 functions in drought and salt stress in transgenic Nicotiana benthamiana through ABA signaling and the modulation of reactive oxygen species production.Plant Cell Physiol. 2014 Dec;55(12):2060-76. doi: 10.1093/pcp/pcu133. Epub 2014 Sep 26. Plant Cell Physiol. 2014. PMID: 25261532
-
Overexpression of a novel SbMYB15 from Salicornia brachiata confers salinity and dehydration tolerance by reduced oxidative damage and improved photosynthesis in transgenic tobacco.Planta. 2015 Dec;242(6):1291-308. doi: 10.1007/s00425-015-2366-5. Epub 2015 Jul 23. Planta. 2015. PMID: 26202734
-
StCaM2, a calcium binding protein, alleviates negative effects of salinity and drought stress in tobacco.Plant Mol Biol. 2021 May;106(1-2):85-108. doi: 10.1007/s11103-021-01131-1. Epub 2021 Feb 24. Plant Mol Biol. 2021. PMID: 33629224
-
A Wheat R2R3-type MYB Transcription Factor TaODORANT1 Positively Regulates Drought and Salt Stress Responses in Transgenic Tobacco Plants.Front Plant Sci. 2017 Aug 8;8:1374. doi: 10.3389/fpls.2017.01374. eCollection 2017. Front Plant Sci. 2017. PMID: 28848578 Free PMC article.
Cited by
-
Abiotic Stresses in Plants and Their Markers: A Practice View of Plant Stress Responses and Programmed Cell Death Mechanisms.Plants (Basel). 2022 Apr 19;11(9):1100. doi: 10.3390/plants11091100. Plants (Basel). 2022. PMID: 35567101 Free PMC article. Review.
-
The Role of Polyphenols in Abiotic Stress Tolerance and Their Antioxidant Properties to Scavenge Reactive Oxygen Species and Free Radicals.Antioxidants (Basel). 2025 Jan 10;14(1):74. doi: 10.3390/antiox14010074. Antioxidants (Basel). 2025. PMID: 39857408 Free PMC article. Review.
-
Molecular insights and omics-based understanding of plant-microbe interactions under drought stress.World J Microbiol Biotechnol. 2023 Dec 18;40(2):42. doi: 10.1007/s11274-023-03837-4. World J Microbiol Biotechnol. 2023. PMID: 38105277 Review.
-
CRISPRi-Mediated Down-Regulation of the Cinnamate-4-Hydroxylase (C4H) Gene Enhances the Flavonoid Biosynthesis in Nicotiana tabacum.Biology (Basel). 2022 Jul 27;11(8):1127. doi: 10.3390/biology11081127. Biology (Basel). 2022. PMID: 36009753 Free PMC article.
-
Functional characterization of 5'-regulatory region of flavonoid 3',5'-hydroxylase-1 gene of banana plants.Protoplasma. 2023 Mar;260(2):391-403. doi: 10.1007/s00709-022-01785-8. Epub 2022 Jun 21. Protoplasma. 2023. PMID: 35727420
References
-
- Ashraf M., Harris P. J. C. (2004). Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 166 3–16. 10.1016/j.plantsci.2003.10.024 - DOI
LinkOut - more resources
Full Text Sources