

The contribution of astrocytes to the neuroinflammatory response in multiple sclerosis and experimental autoimmune encephalomyelitis
- PMID: 30847559
- PMCID: PMC6483860
- DOI: 10.1007/s00401-019-01980-7
The contribution of astrocytes to the neuroinflammatory response in multiple sclerosis and experimental autoimmune encephalomyelitis
Abstract
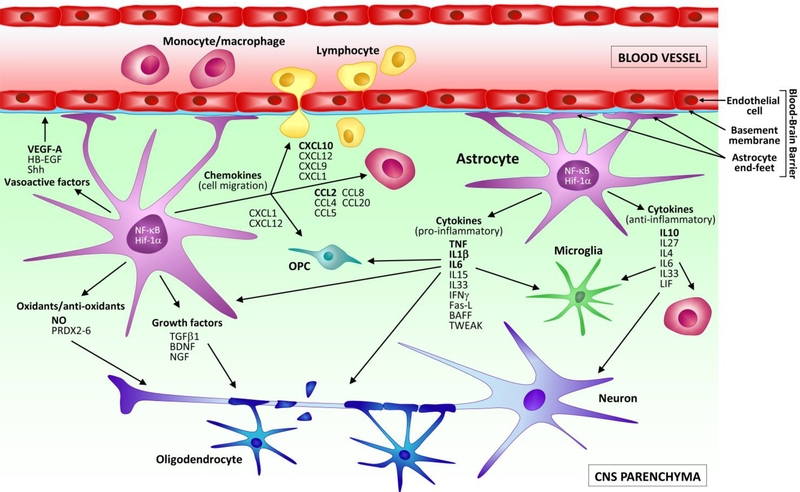
Neuroinflammation is the coordinated response of the central nervous system (CNS) to threats to its integrity posed by a variety of conditions, including autoimmunity, pathogens and trauma. Activated astrocytes, in concert with other cellular elements of the CNS and immune system, are important players in the modulation of the neuroinflammatory response. During neurological disease, they produce and respond to cellular signals that often lead to dichotomous processes, which can promote further damage or contribute to repair. This occurs also in multiple sclerosis (MS), where astrocytes are now recognized as key components of its immunopathology. Evidence supporting this role has emerged not only from studies in MS patients, but also from animal models, among which the experimental autoimmune encephalomyelitis (EAE) model has proved especially instrumental. Based on this premise, the purpose of the present review is to summarize the current knowledge of astrocyte behavior in MS and EAE. Following a brief description of the pathological characteristics of the two diseases and the main functional roles of astrocytes in CNS physiology, we will delve into the specific responses of this cell population, analyzing MS and EAE in parallel. We will define the temporal and anatomical profile of astroglial activation, then focus on key processes they participate in. These include: (1) production and response to soluble mediators (e.g., cytokines and chemokines), (2) regulation of oxidative stress, and (3) maintenance of BBB integrity and function. Finally, we will review the state of the art on the available methods to measure astroglial activation in vivo in MS patients, and how this could be exploited to optimize diagnosis, prognosis and treatment decisions. Ultimately, we believe that integrating the knowledge obtained from studies in MS and EAE may help not only better understand the pathophysiology of MS, but also uncover new signals to be targeted for therapeutic intervention.
Keywords: Astroglia; Demyelinating disorder; Experimental autoimmune encephalomyelitis; Multiple sclerosis; Neuroimmune disease; Neuroinflammation.
Conflict of interest statement
Figures
References
-
- Agrawal S, Anderson P, Durbeej M, van Rooijen N, Ivars F, Opdenakker G, Sorokin LM (2006) Dystroglycan is selectively cleaved at the parenchymal basement membrane at sites of leukocyte extravasation in experimental autoimmune encephalomyelitis. J Exp Med 203: 1007–1019 Doi 10.1084/jem.20051342 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
