

Specific and Selective Inhibitors of Proprotein Convertases Engineered by Transferring Serpin B8 Reactive-Site and Exosite Determinants of Reactivity to the Serpin α1PDX
- PMID: 30848586
- PMCID: PMC6469502
- DOI: 10.1021/acs.biochem.8b01295
Specific and Selective Inhibitors of Proprotein Convertases Engineered by Transferring Serpin B8 Reactive-Site and Exosite Determinants of Reactivity to the Serpin α1PDX
Abstract
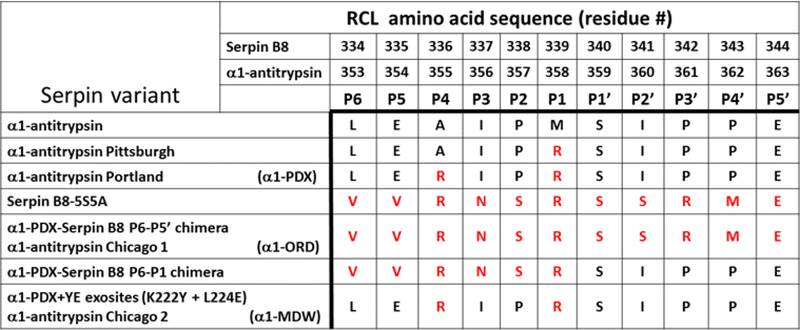
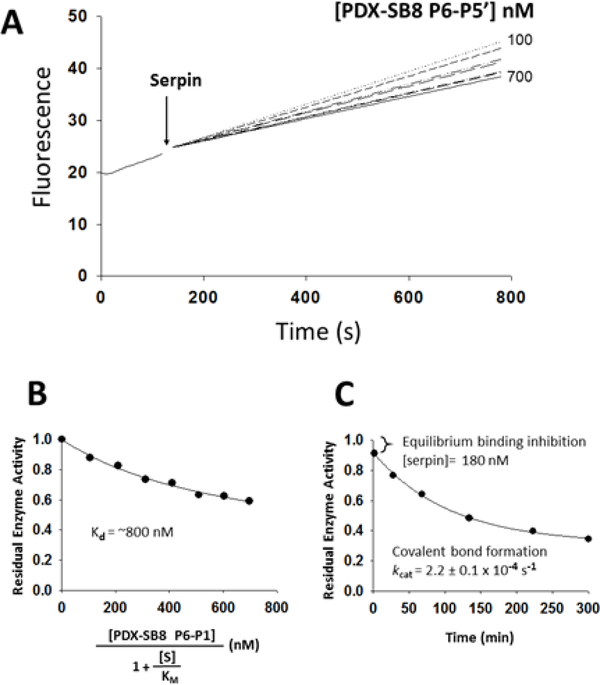
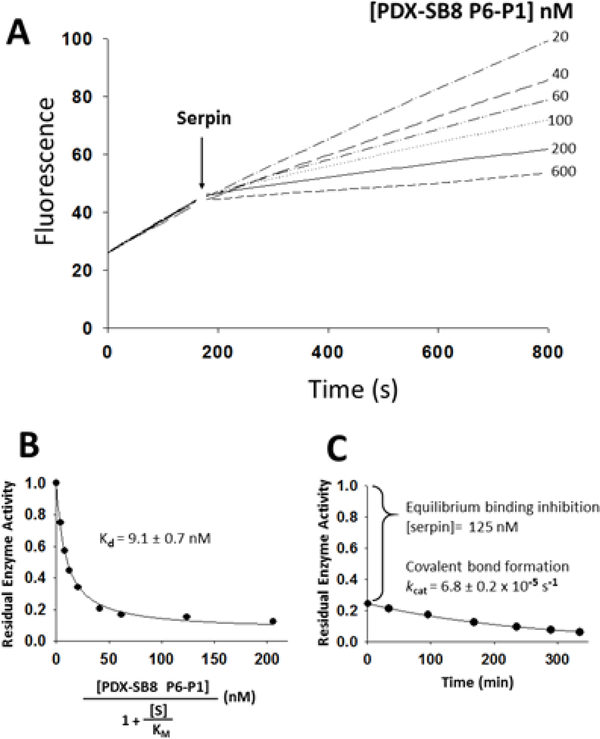
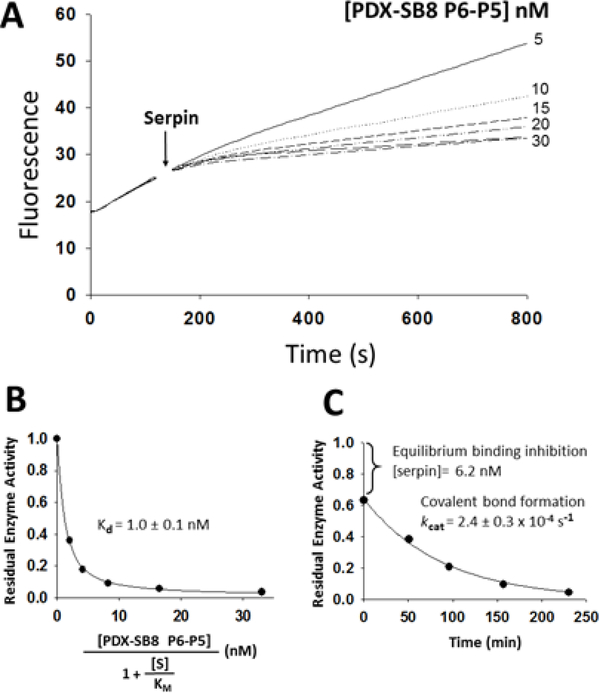
The molecular determinants of substrate specificity and selectivity in the proprotein convertase (PC) family of proteases are poorly understood. Here we demonstrate that the natural serpin family inhibitor, serpin B8, is a specific and selective inhibitor of furin relative to the other PCs of the constitutive protein secretion pathway, PC4, PC5, PACE4, and PC7 (PC4-PC7, respectively), and identify reactive-site (P6-P5' residues) and exosite elements of the serpin that contribute to this specificity and selectivity through studies of chimeras of serpin B8 and α1PDX, an engineered serpin inhibitor of furin. Kinetic studies revealed that the specificity and selectivity of the serpin chimeras for inhibiting PCs were determined by P6-P5 and P3-P2 residue-dependent recognition of the P4Arg-X-X-P1Arg PC consensus sequence and exosite-dependent recognition of the reactive loop P2' residue of the chimeras by the PCs. Both productive and nonproductive binding of the chimeras to PC4-PC7 but not to furin contributed to a decreased specificity for inhibiting PC4-PC7 and an increased selectivity for inhibiting furin. Molecular dynamics simulations suggested that nonproductive binding of the chimeras to the PCs was correlated with a greater conformational variability of the catalytic sites of PC4-PC7 relative to that of furin. Our findings suggest a new approach for designing selective inhibitors of PCs using α1PDX as a scaffold, as evidenced by our ability to engineer highly specific and selective inhibitors of furin and PC4-PC7.
Figures
Similar articles
-
The Proteolytic Regulation of Virus Cell Entry by Furin and Other Proprotein Convertases.Viruses. 2019 Sep 9;11(9):837. doi: 10.3390/v11090837. Viruses. 2019. PMID: 31505793 Free PMC article. Review.
-
Identification of serpin determinants of specificity and selectivity for furin inhibition through studies of α1PDX (α1-protease inhibitor Portland)-serpin B8 and furin active-site loop chimeras.J Biol Chem. 2013 Jul 26;288(30):21802-14. doi: 10.1074/jbc.M113.462804. Epub 2013 Jun 6. J Biol Chem. 2013. PMID: 23744066 Free PMC article.
-
Comparative study of the binding pockets of mammalian proprotein convertases and its implications for the design of specific small molecule inhibitors.Int J Biol Sci. 2010 Feb 3;6(1):89-95. doi: 10.7150/ijbs.6.89. Int J Biol Sci. 2010. PMID: 20151049 Free PMC article.
-
Engineering of α1-antitrypsin variants with improved specificity for the proprotein convertase furin using site-directed random mutagenesis.Protein Eng Des Sel. 2013 Feb;26(2):123-31. doi: 10.1093/protein/gzs091. Epub 2012 Nov 14. Protein Eng Des Sel. 2013. PMID: 23155057
-
The proprotein convertases furin and PACE4 play a significant role in tumor progression.Mol Carcinog. 2000 Jun;28(2):63-9. Mol Carcinog. 2000. PMID: 10900462 Review.
Cited by
-
Diversity in Proprotein Convertase Reactivity among Human Papillomavirus Types.Viruses. 2023 Dec 26;16(1):39. doi: 10.3390/v16010039. Viruses. 2023. PMID: 38257739 Free PMC article.
-
The Use of Tick Salivary Proteins as Novel Therapeutics.Front Physiol. 2019 Jun 26;10:812. doi: 10.3389/fphys.2019.00812. eCollection 2019. Front Physiol. 2019. PMID: 31297067 Free PMC article. Review.
-
Restriction of Viral Glycoprotein Maturation by Cellular Protease Inhibitors.Viruses. 2024 Feb 22;16(3):332. doi: 10.3390/v16030332. Viruses. 2024. PMID: 38543698 Free PMC article. Review.
-
The Proteolytic Regulation of Virus Cell Entry by Furin and Other Proprotein Convertases.Viruses. 2019 Sep 9;11(9):837. doi: 10.3390/v11090837. Viruses. 2019. PMID: 31505793 Free PMC article. Review.
-
Engineering the serpin α1 -antitrypsin: A diversity of goals and techniques.Protein Sci. 2020 Apr;29(4):856-871. doi: 10.1002/pro.3794. Epub 2019 Dec 9. Protein Sci. 2020. PMID: 31774589 Free PMC article. Review.
References
-
- Seidah NG, and Prat A (2012) The biology and therapeutic targeting of the proprotein convertases. Nat. Rev. Drug Discov 11: 367–383 - PubMed
-
- Seidah NG, Mayer G, Zaid A, Rousselet E, Nassoury N, Poirier S, Essalmani R, and Prat A (2008) The activation and physiological functions of the proprotein convertases. Int. J. Biochem. Cell Biol 40: 1111–1125 - PubMed
-
- Shiryaev SA, Remacle AG, Ratnikov BI, Nelson NA, Savinov AY, Wei G, Bottini M, Rega MF, Parent A, Desjardins R, Fugere M, Day R, Sabet M, Pellecchia M, Liddington RC, Smith JW, Mustelin T, Guiney DG, Lebl M, and Strongin AY (2007) Targeting host cell furin proprotein convertases as a therapeutic strategy against bacterial toxins and viral pathogens. J. Biol. Chem 282: 20847–20853 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
