

Higher-Order Thalamic Circuits Channel Parallel Streams of Visual Information in Mice
- PMID: 30850257
- PMCID: PMC8638696
- DOI: 10.1016/j.neuron.2019.02.010
Higher-Order Thalamic Circuits Channel Parallel Streams of Visual Information in Mice
Abstract
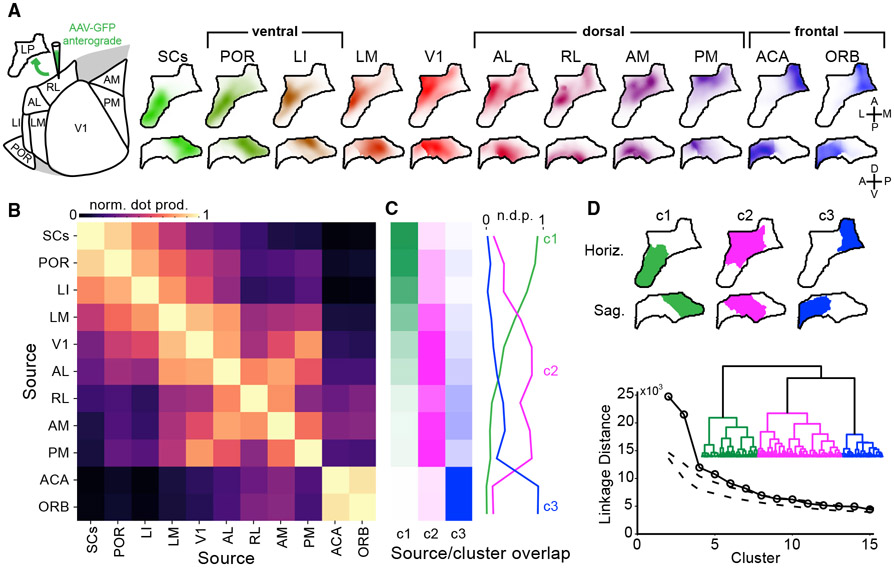
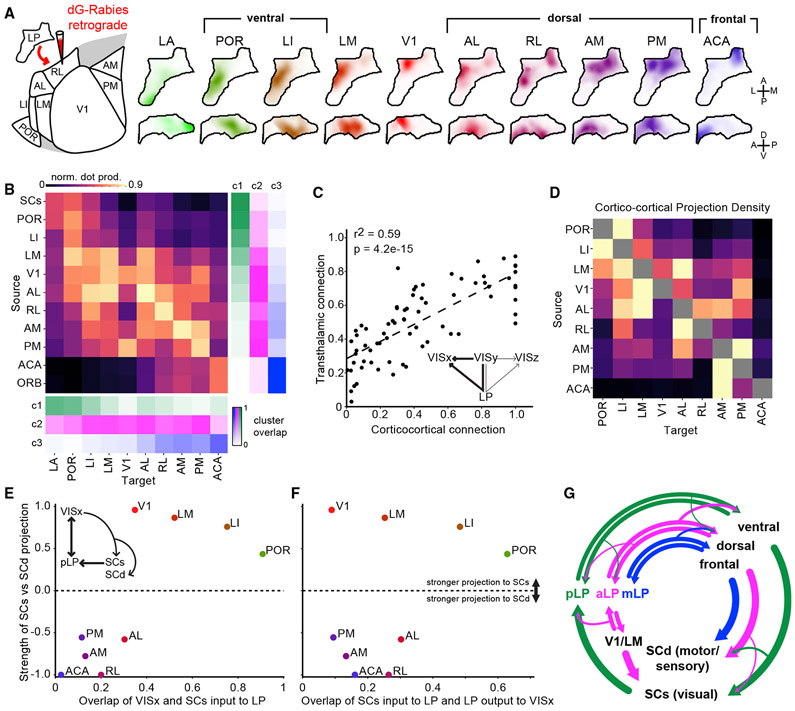
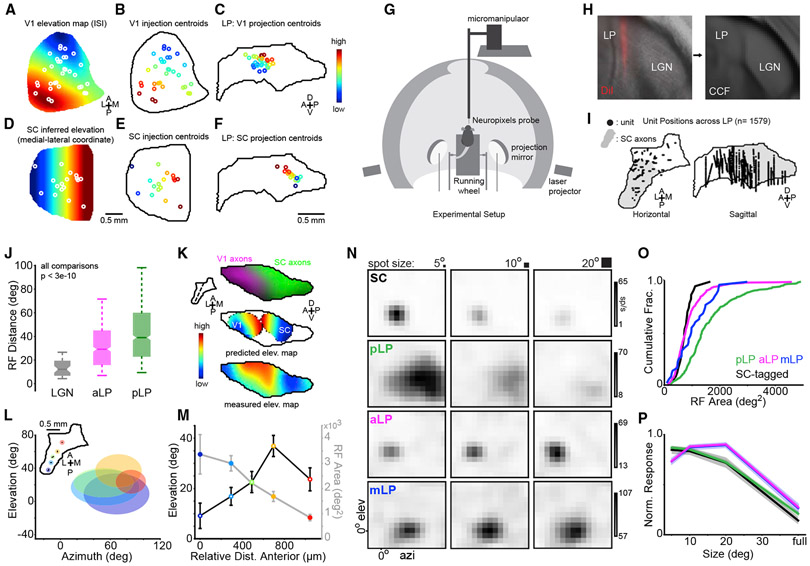
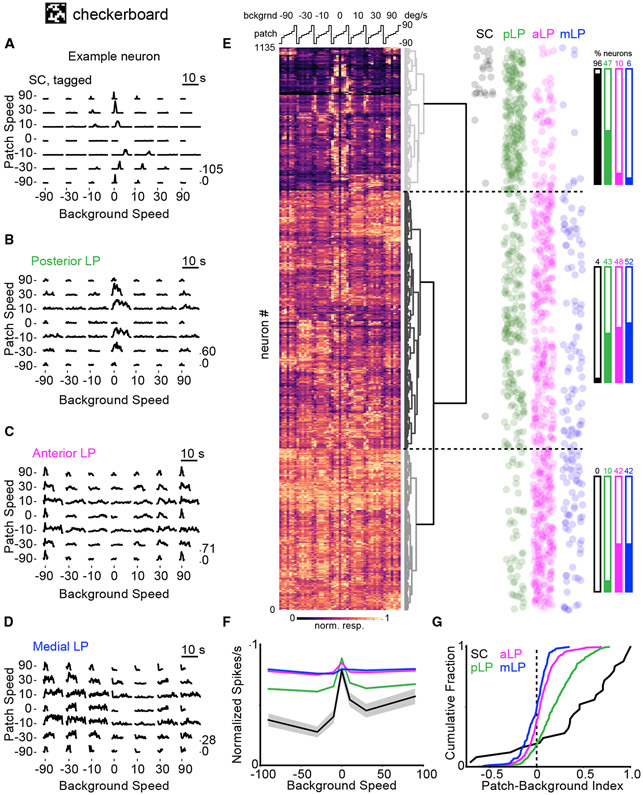
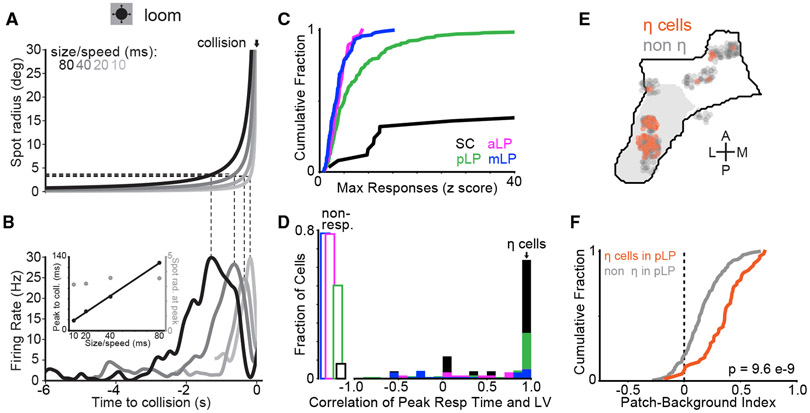
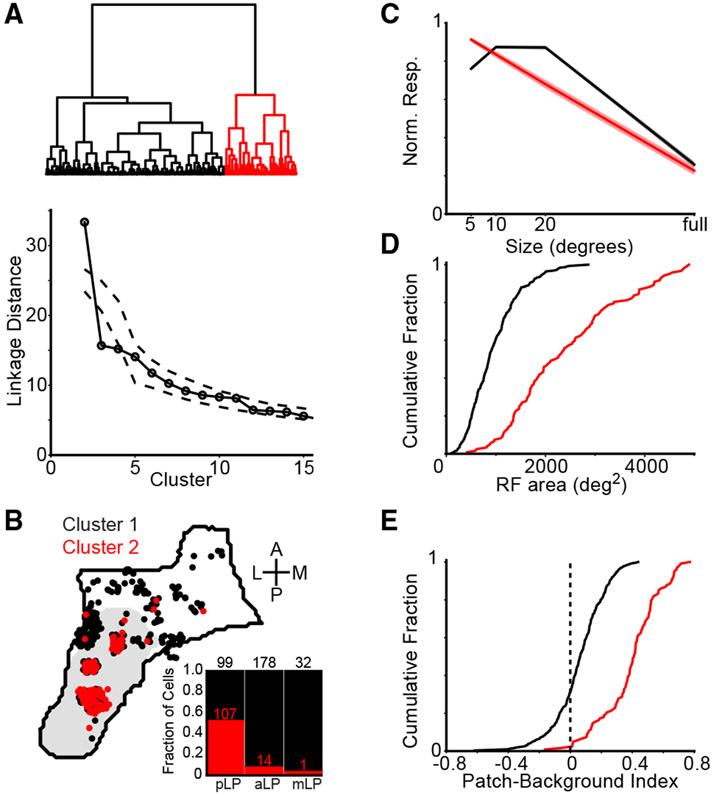
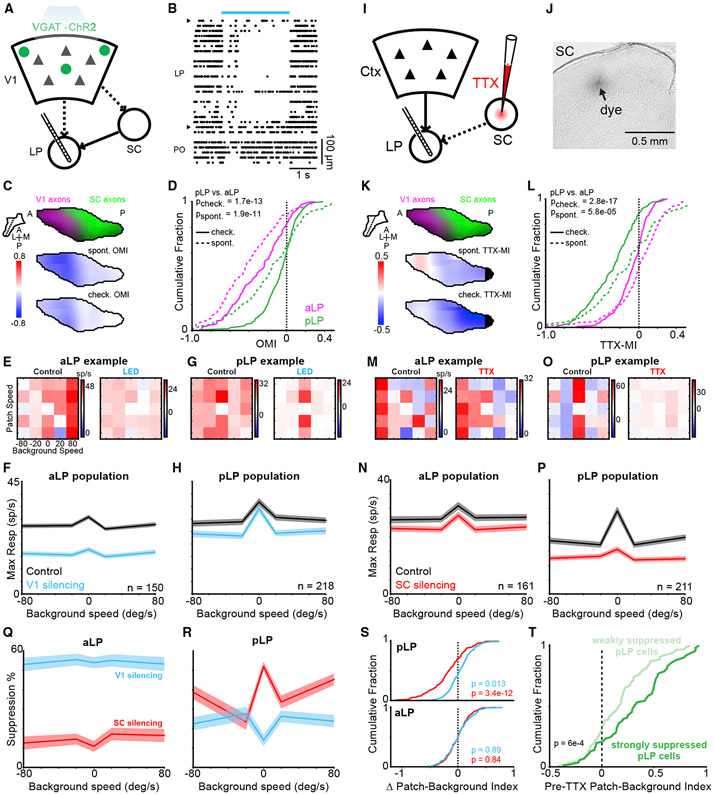
Higher-order thalamic nuclei, such as the visual pulvinar, play essential roles in cortical function by connecting functionally related cortical and subcortical brain regions. A coherent framework describing pulvinar function remains elusive because of its anatomical complexity and involvement in diverse cognitive processes. We combined large-scale anatomical circuit mapping with high-density electrophysiological recordings to dissect a homolog of the pulvinar in mice, the lateral posterior thalamic nucleus (LP). We define three broad LP subregions based on correspondence between connectivity and functional properties. These subregions form corticothalamic loops biased toward ventral or dorsal stream cortical areas and contain separate representations of visual space. Silencing the visual cortex or superior colliculus revealed that they drive visual tuning properties in separate LP subregions. Thus, by specifying the driving input sources, functional properties, and downstream targets of LP circuits, our data provide a roadmap for understanding the mechanisms of higher-order thalamic function in vision.
Keywords: lateral posterior thalamic nucleus; pulvinar; superior colliculus; thalamus; visual cortex.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
References
-
- Baldwin MKL, Balaram P, and Kaas JH (2017). The evolution and functions of nuclei of the visual pulvinar in primates. J. Comp. Neurol 525, 3207–3226. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
