

Diversifying the structure of zinc finger nucleases for high-precision genome editing
- PMID: 30850604
- PMCID: PMC6408524
- DOI: 10.1038/s41467-019-08867-x
Diversifying the structure of zinc finger nucleases for high-precision genome editing
Abstract
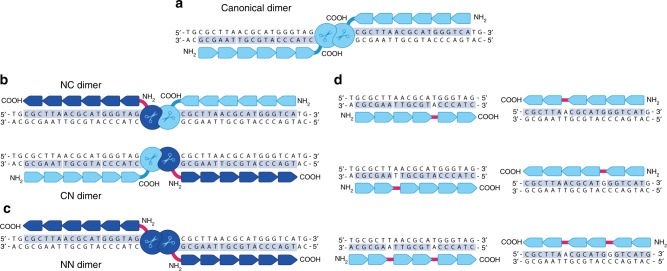
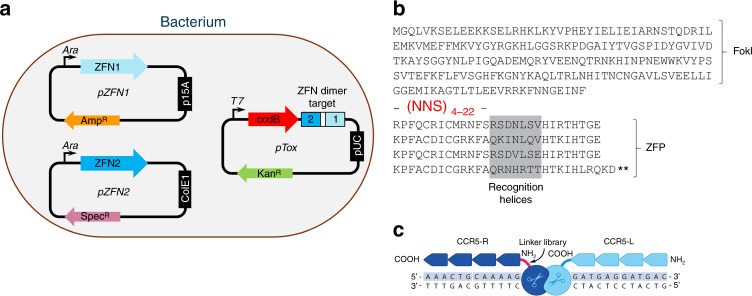
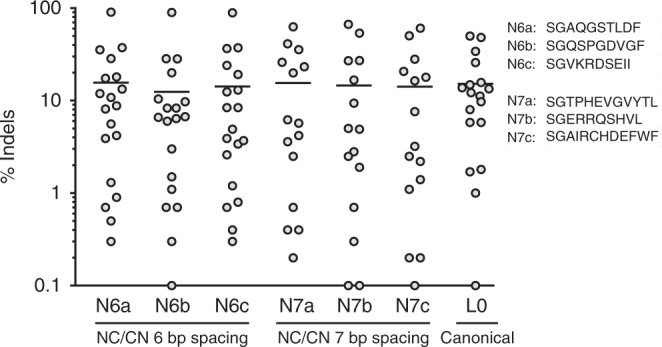
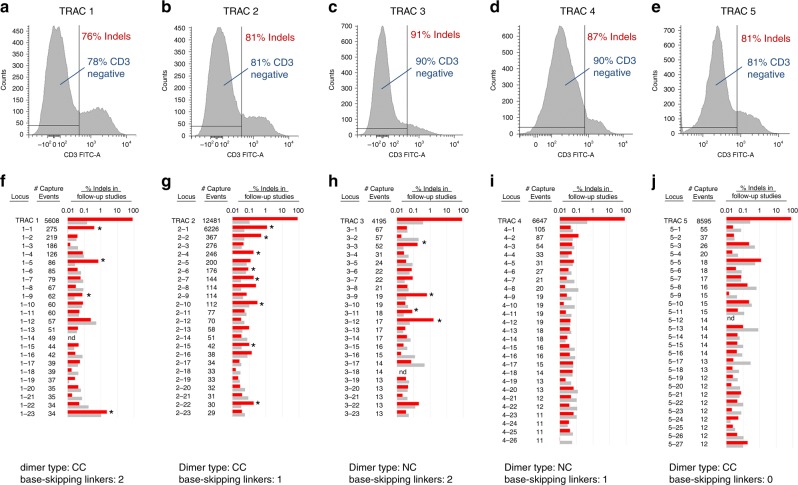
Genome editing for therapeutic applications often requires cleavage within a narrow sequence window. Here, to enable such high-precision targeting with zinc-finger nucleases (ZFNs), we have developed an expanded set of architectures that collectively increase the configurational options available for design by a factor of 64. These new architectures feature the functional attachment of the FokI cleavage domain to the amino terminus of one or both zinc-finger proteins (ZFPs) in the ZFN dimer, as well as the option to skip bases between the target triplets of otherwise adjacent fingers in each zinc-finger array. Using our new architectures, we demonstrate targeting of an arbitrarily chosen 28 bp genomic locus at a density that approaches 1.0 (i.e., efficient ZFNs available for targeting almost every base step). We show that these new architectures may be used for targeting three loci of therapeutic significance with a high degree of precision, efficiency, and specificity.
Conflict of interest statement
All authors are current or former full-time employees of Sangamo Therapeutics, Inc.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
