

The Cargo Receptor NDP52 Initiates Selective Autophagy by Recruiting the ULK Complex to Cytosol-Invading Bacteria
- PMID: 30853402
- PMCID: PMC6477152
- DOI: 10.1016/j.molcel.2019.01.041
The Cargo Receptor NDP52 Initiates Selective Autophagy by Recruiting the ULK Complex to Cytosol-Invading Bacteria
Abstract
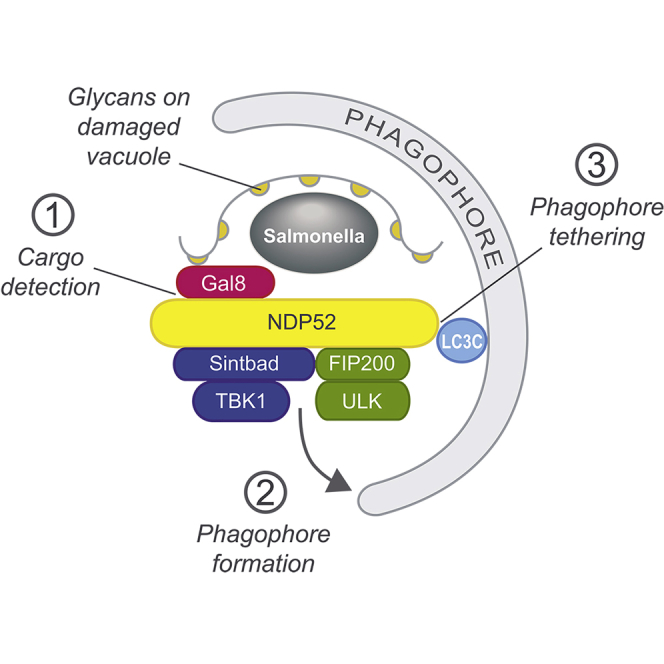
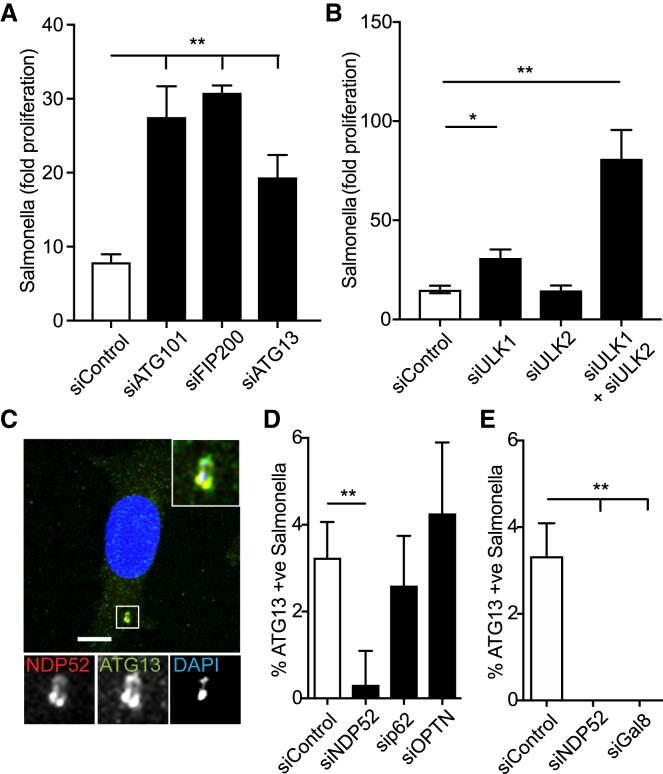
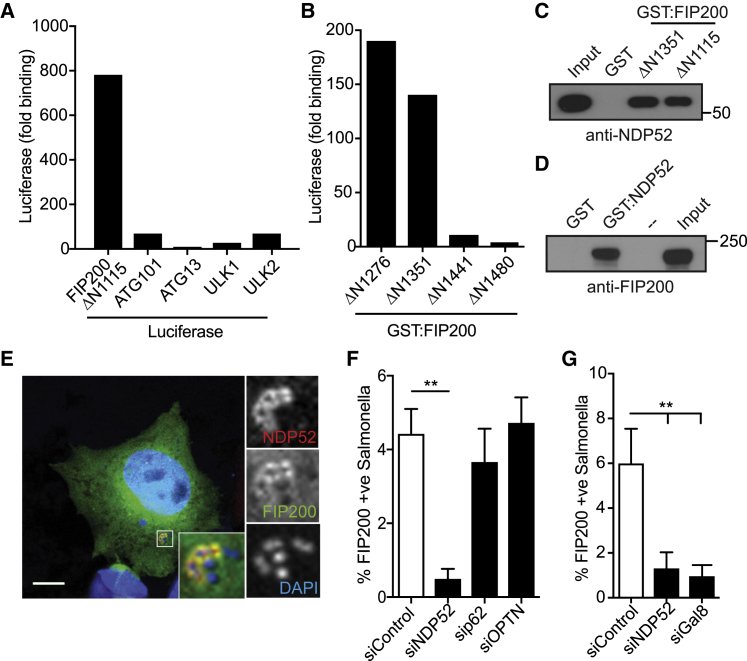
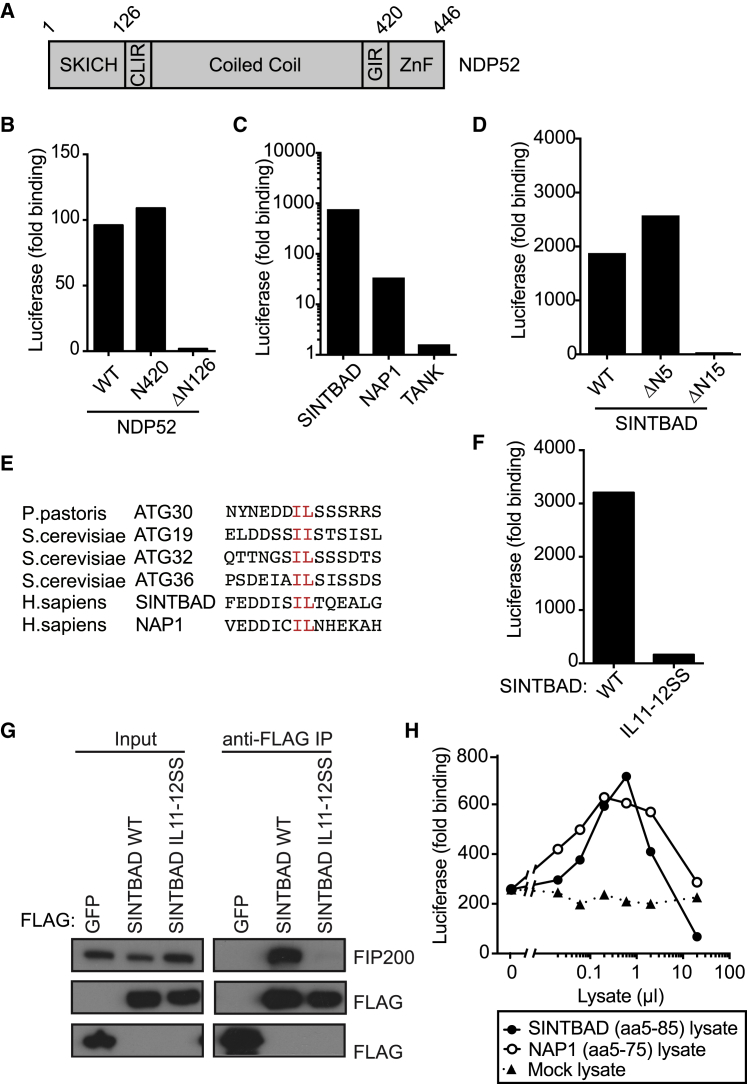
Xenophagy, a selective autophagy pathway that protects the cytosol against bacterial invasion, relies on cargo receptors that juxtapose bacteria and phagophore membranes. Whether phagophores are recruited from a constitutive pool or are generated de novo at prospective cargo remains unknown. Phagophore formation in situ would require recruitment of the upstream autophagy machinery to prospective cargo. Here, we show that, essential for anti-bacterial autophagy, the cargo receptor NDP52 forms a trimeric complex with FIP200 and SINTBAD/NAP1, which are subunits of the autophagy-initiating ULK and the TBK1 kinase complex, respectively. FIP200 and SINTBAD/NAP1 are each recruited independently to bacteria via NDP52, as revealed by selective point mutations in their respective binding sites, but only in their combined presence does xenophagy proceed. Such recruitment of the upstream autophagy machinery by NDP52 reveals how detection of cargo-associated "eat me" signals, induction of autophagy, and juxtaposition of cargo and phagophores are integrated in higher eukaryotes.
Keywords: FIP200; NDP52; Salmonella enterica; TBK1; ULK; cargo receptor; galectin-8; selective autophagy; xenophagy.
Copyright © 2019 MRC Laboratory of Molecular Biology. Published by Elsevier Inc. All rights reserved.
Figures
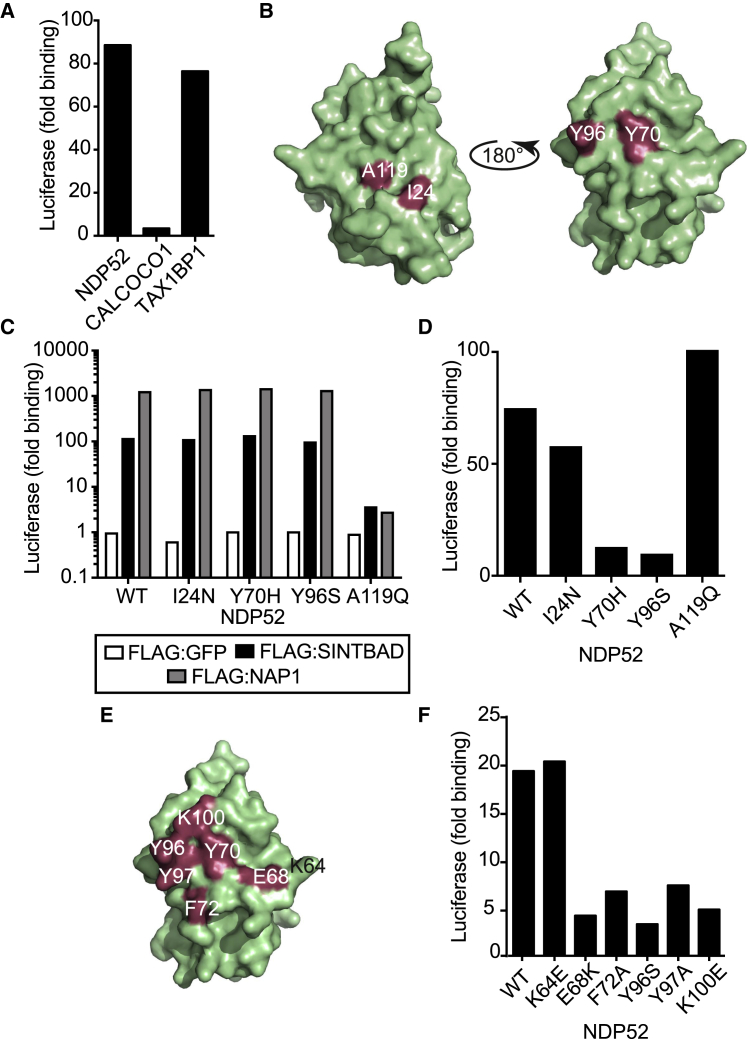
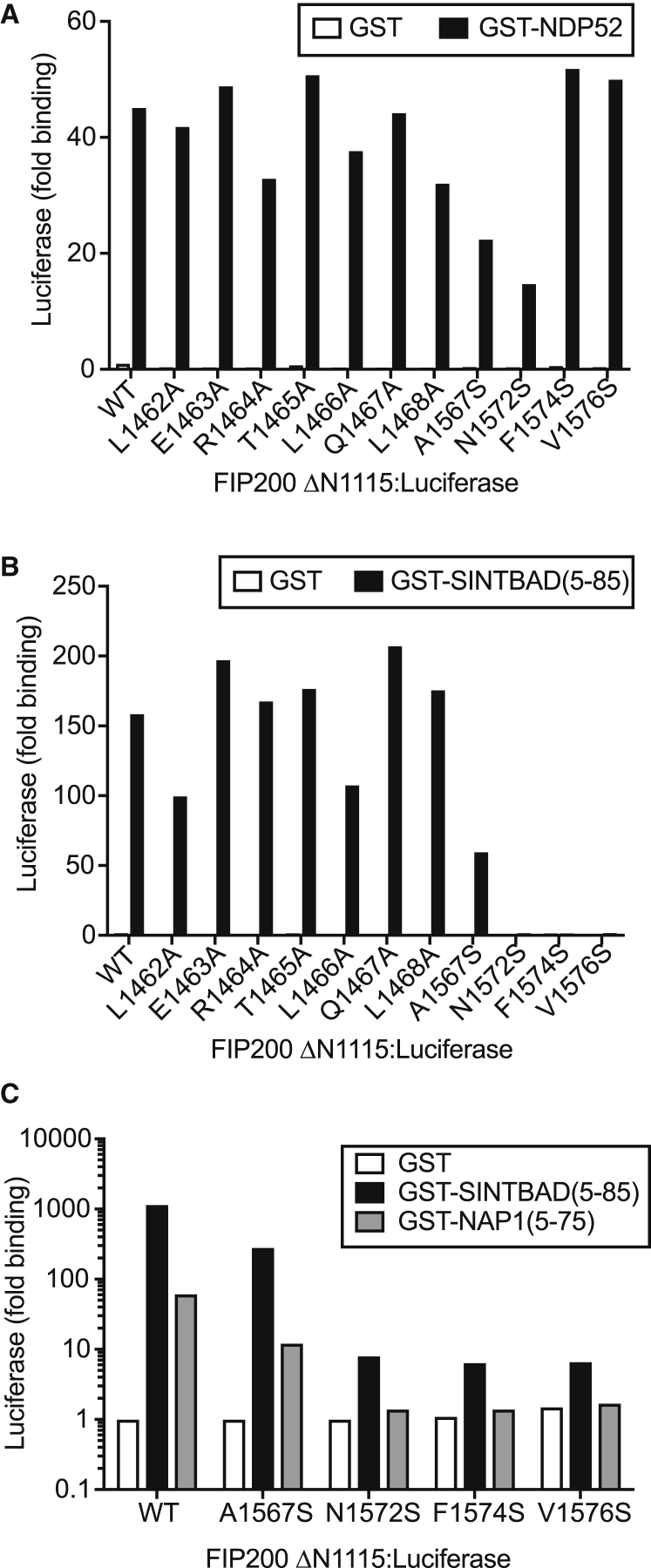
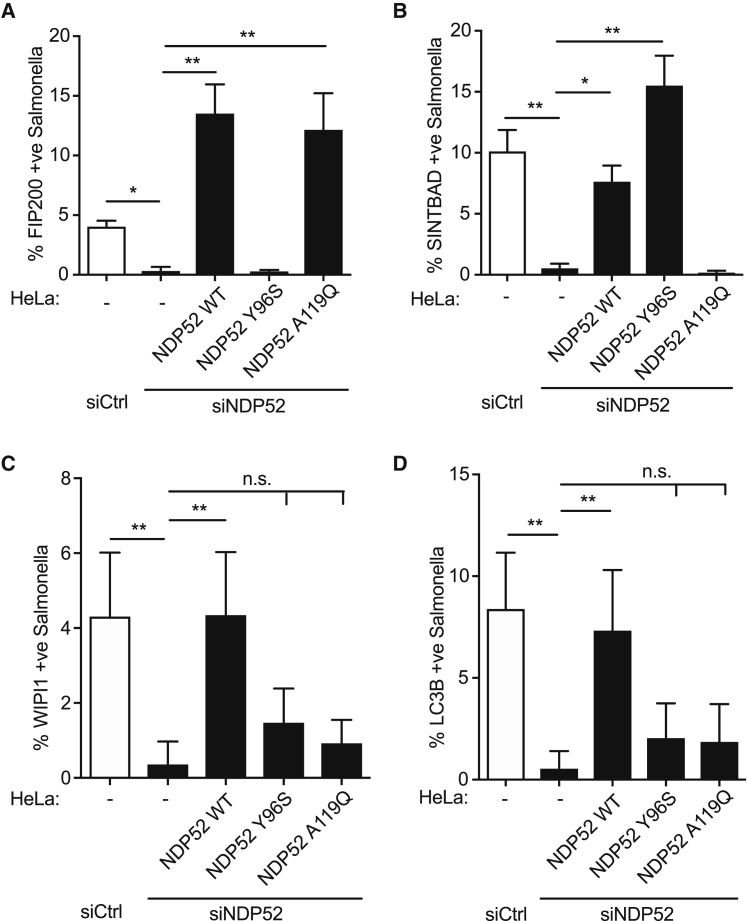
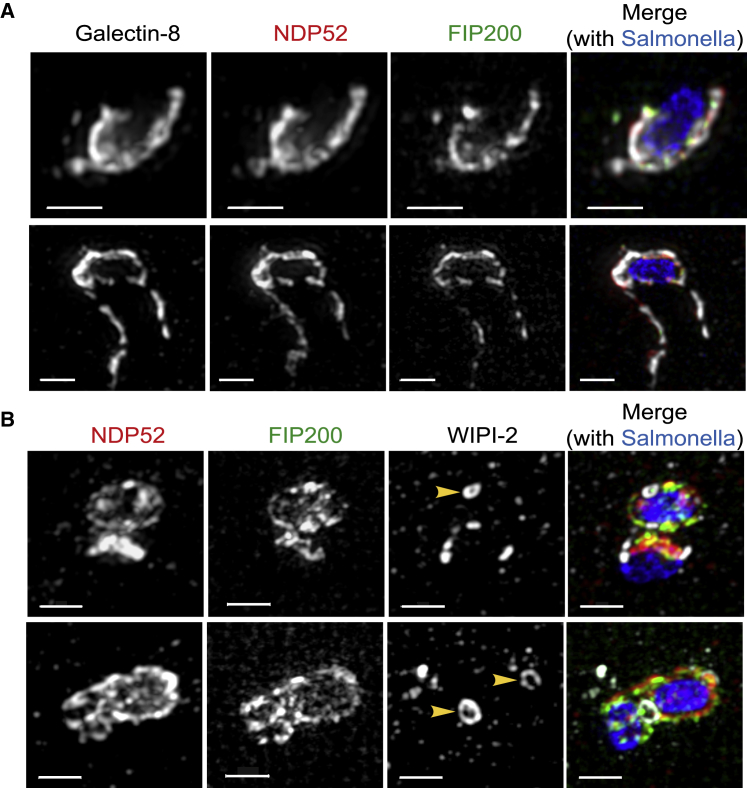
Comment in
-
CALCOCO2/NDP52 initiates selective autophagy through recruitment of ULK and TBK1 kinase complexes.Autophagy. 2019 Sep;15(9):1655-1656. doi: 10.1080/15548627.2019.1628548. Epub 2019 Jul 1. Autophagy. 2019. PMID: 31258038 Free PMC article.
References
-
- Bouwmeester T., Bauch A., Ruffner H., Angrand P.-O., Bergamini G., Croughton K., Cruciat C., Eberhard D., Gagneur J., Ghidelli S. A physical and functional map of the human TNF-α/NF-κ B signal transduction pathway. Nat. Cell Biol. 2004;6:97–105. - PubMed
-
- Boyle K.B., Randow F. The role of ‘eat-me’ signals and autophagy cargo receptors in innate immunity. Curr. Opin. Microbiol. 2013;16:339–348. - PubMed
-
- Dikic I., Elazar Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Biol. 2018;19:349–364. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
