

Production of YP170 Vitellogenins Promotes Intestinal Senescence in Caenorhabditis elegans
- PMID: 30854561
- PMCID: PMC6625598
- DOI: 10.1093/gerona/glz067
Production of YP170 Vitellogenins Promotes Intestinal Senescence in Caenorhabditis elegans
Abstract
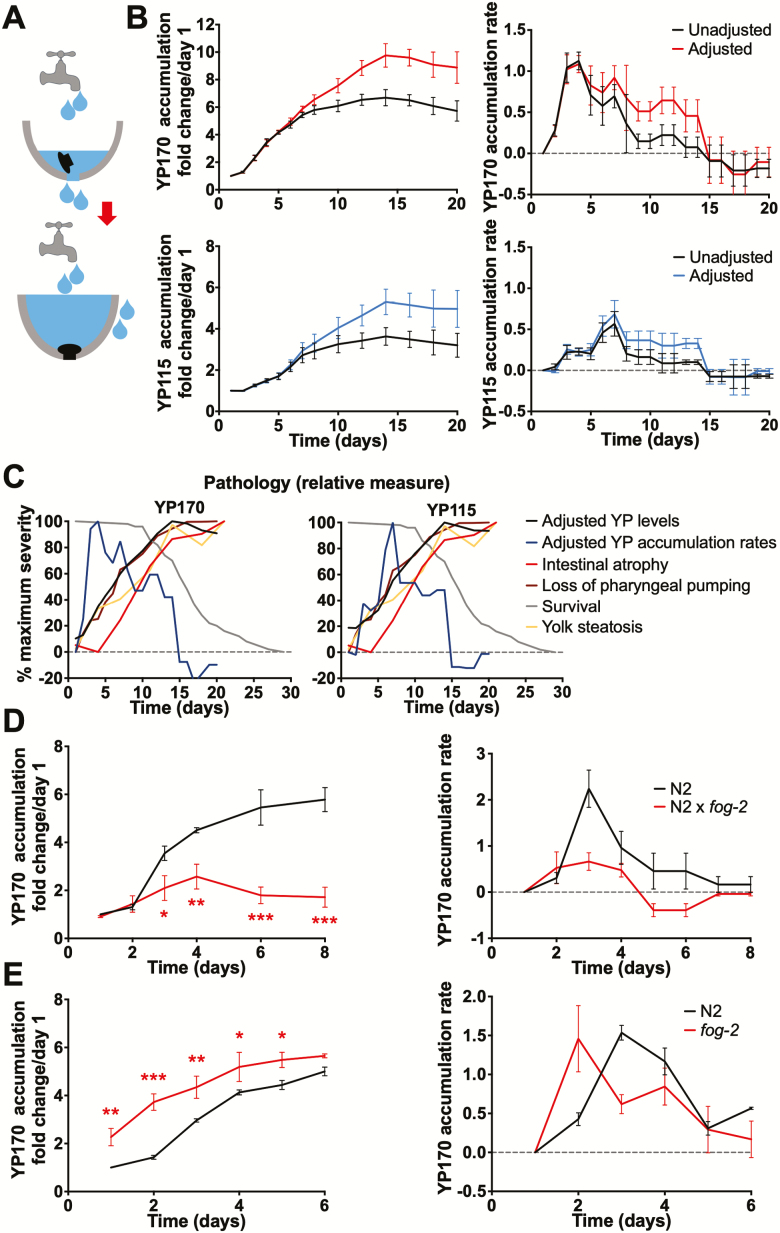
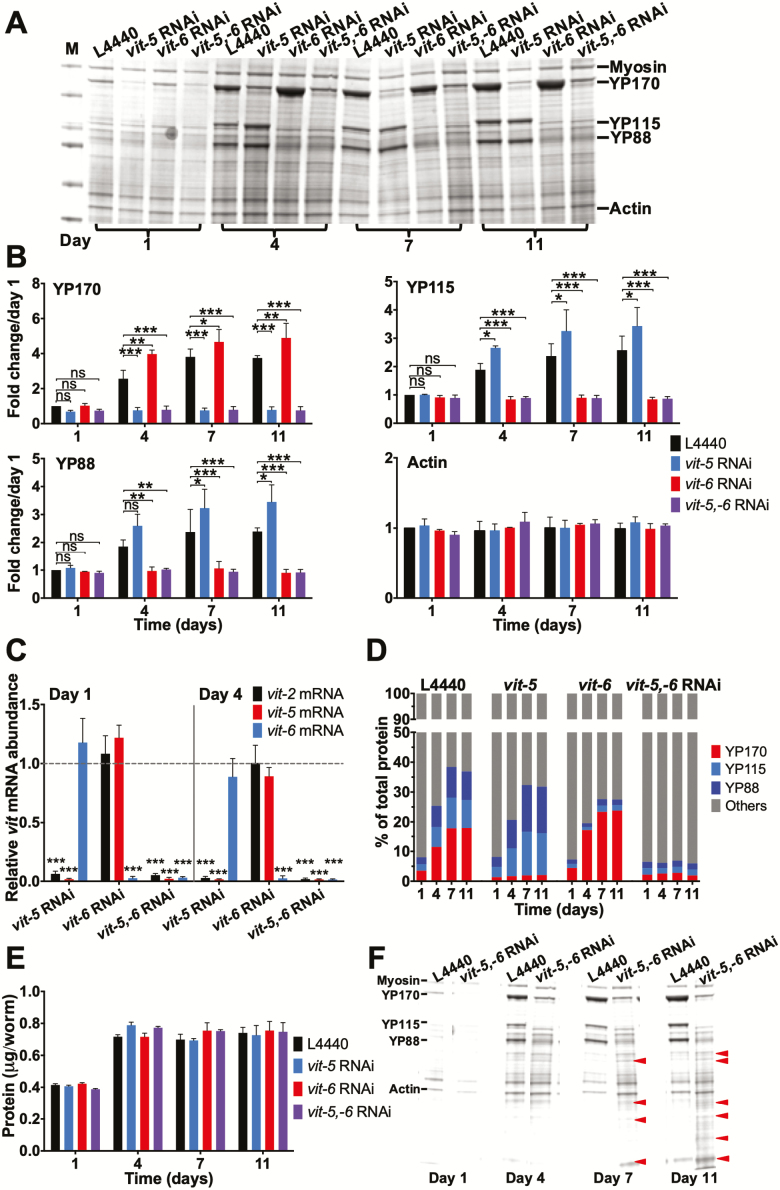
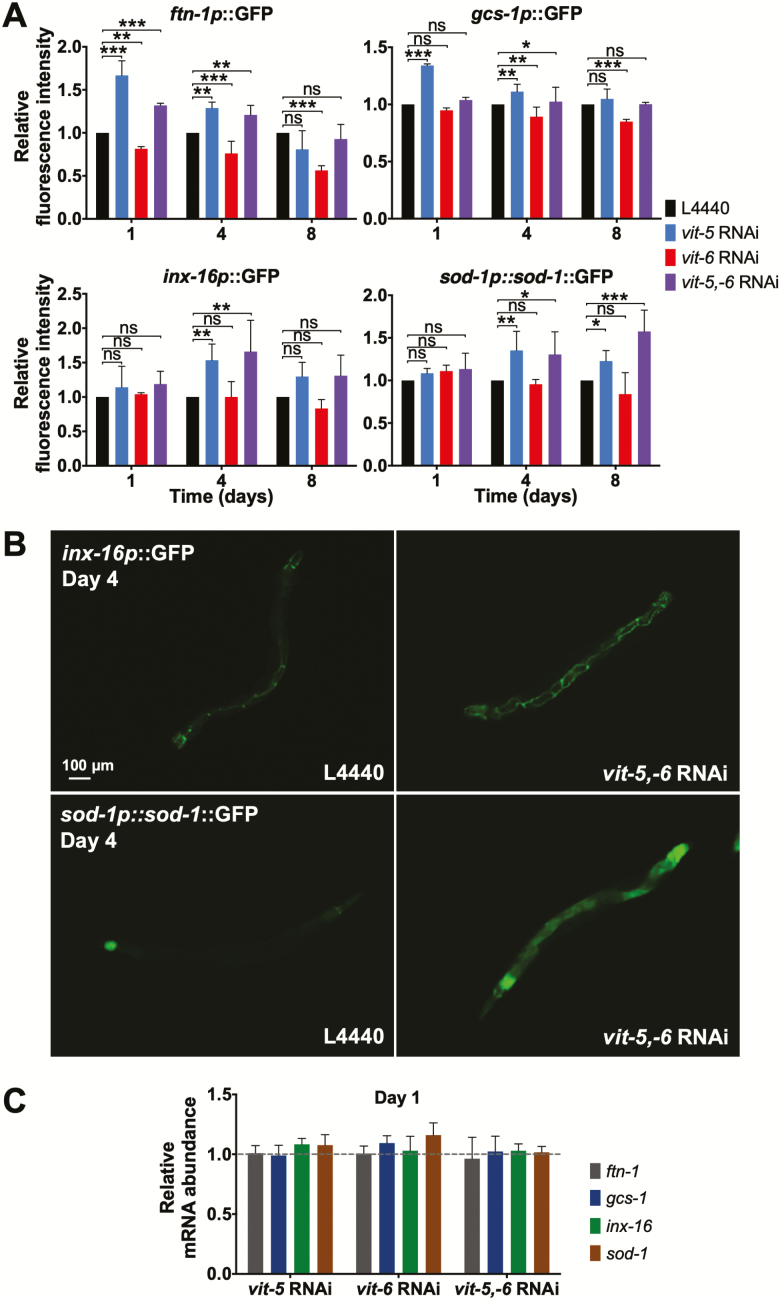
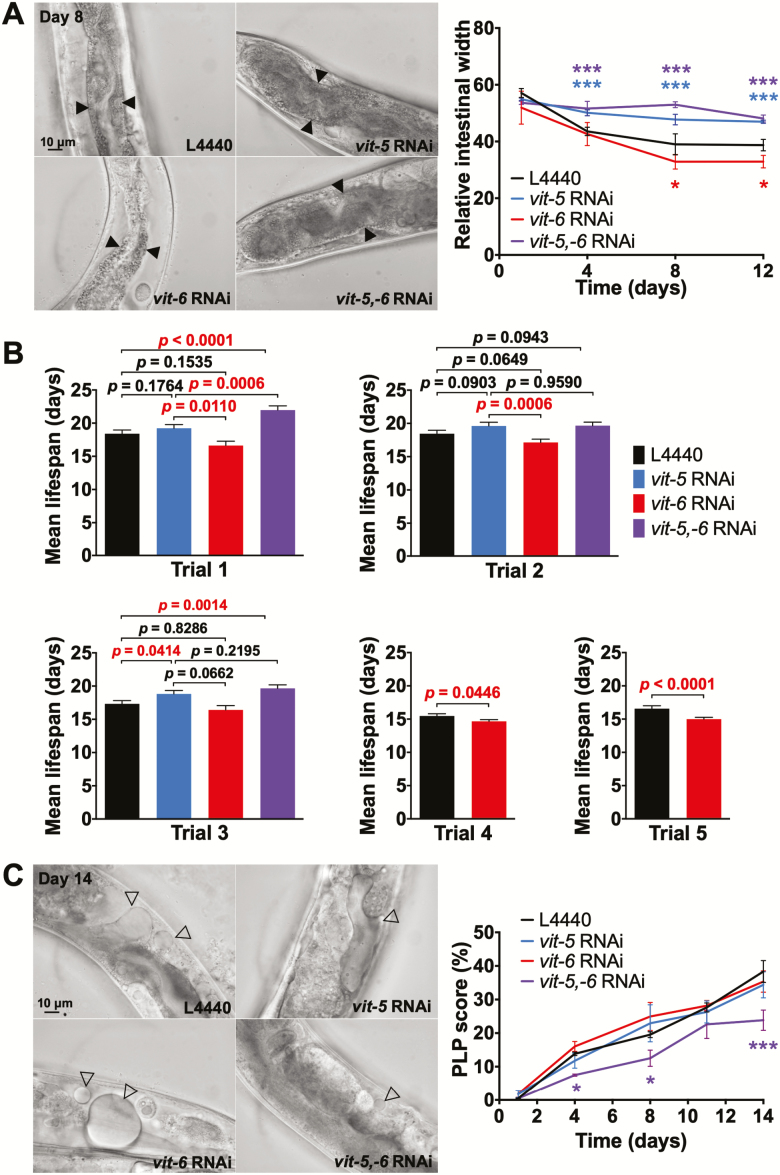
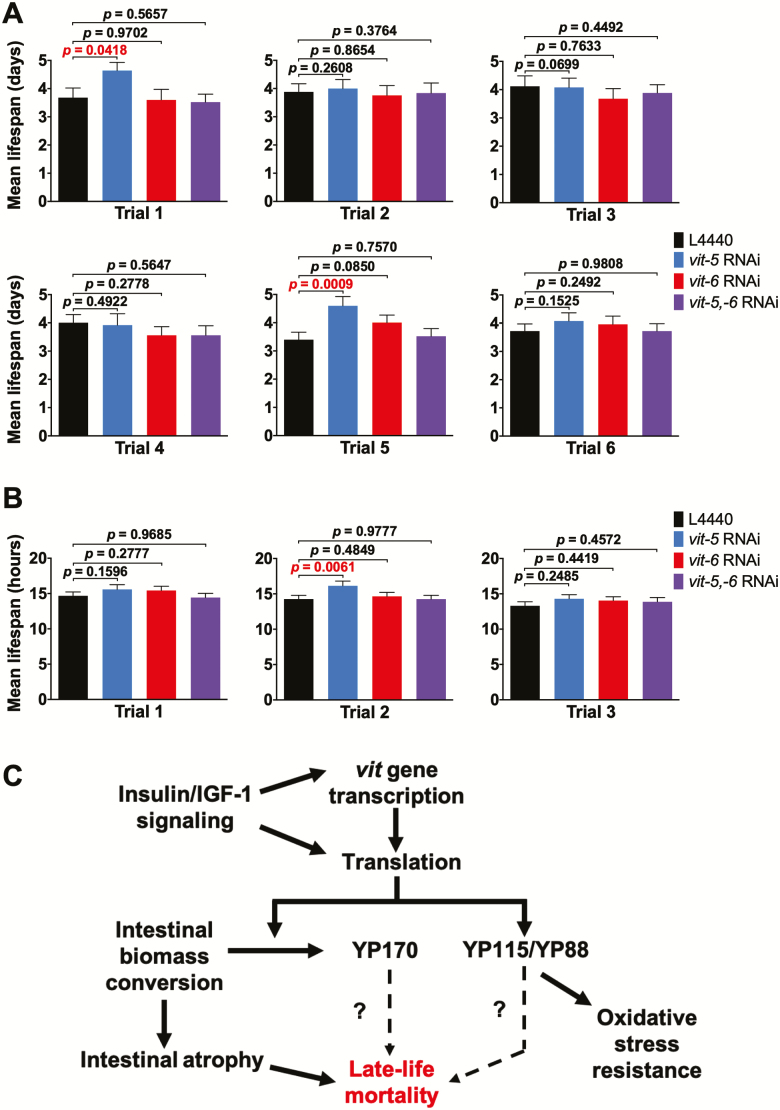
During aging, etiologies of senescence cause multiple pathologies, leading to morbidity and death. To understand aging requires identification of these etiologies. For example, Caenorhabditis elegans hermaphrodites consume their own intestinal biomass to support yolk production, which in later life drives intestinal atrophy and ectopic yolk deposition. Yolk proteins (YPs; vitellogenins) exist as three abundant species: YP170, derived from vit-1-vit-5; and YP115 and YP88, derived from vit-6. Here, we show that inhibiting YP170 synthesis leads to a reciprocal increase in YP115/YP88 levels and vice versa, an effect involving posttranscriptional mechanisms. Inhibiting YP170 production alone, despite increasing YP115/YP88 synthesis, reduces intestinal atrophy as much as inhibition of all YP synthesis, which increases life span. By contrast, inhibiting YP115/YP88 production alone accelerates intestinal atrophy and reduces life span, an effect that is dependent on increased YP170 production. Thus, despite copious abundance of both YP170 and YP115/YP88, only YP170 production is coupled to intestinal atrophy and shortened life span. In addition, increasing levels of YP115/YP88 but not of YP170 increases resistance to oxidative stress; thus, longevity resulting from reduced vitellogenin synthesis is not attributable to oxidative stress resistance.
Keywords: Caenorhabditis elegans; Age-related pathology; Aging; Animal model; Yolk.
© The Author(s) 2019. Published by Oxford University Press on behalf of The Gerontological Society of America.
Figures
References
-
- Williams GC. Pleiotropy, natural selection and the evolution of senescence. Evolution. 1957;11:398–411. doi: 10.2307/2406060 - DOI
