

Early life stress delays hippocampal development and diminishes the adult stem cell pool in mice
- PMID: 30858462
- PMCID: PMC6412041
- DOI: 10.1038/s41598-019-40868-0
Early life stress delays hippocampal development and diminishes the adult stem cell pool in mice
Abstract
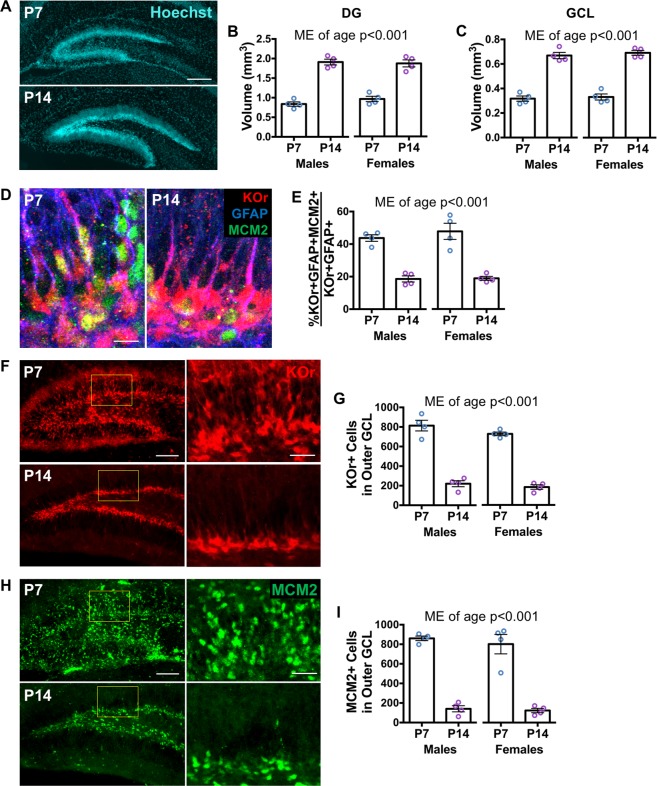
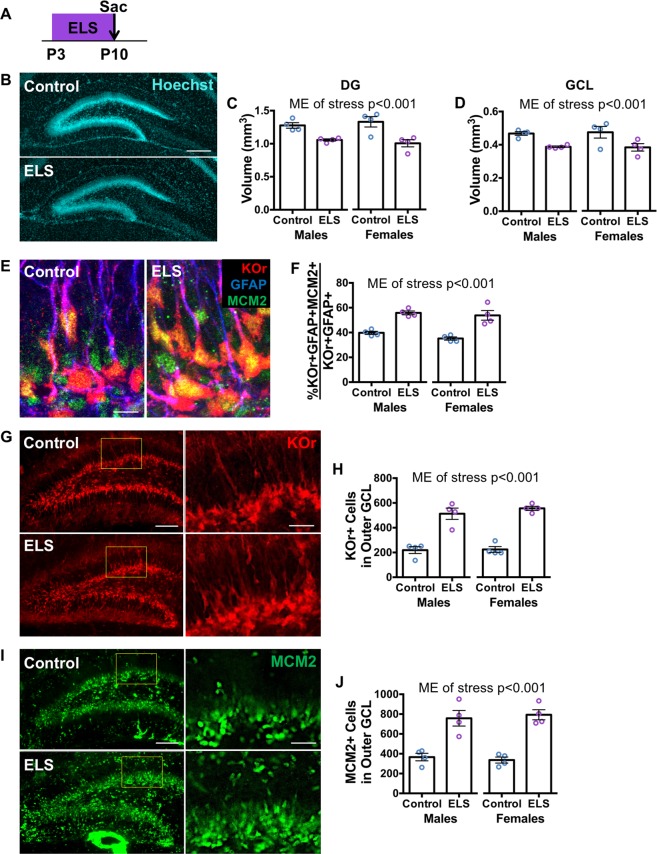
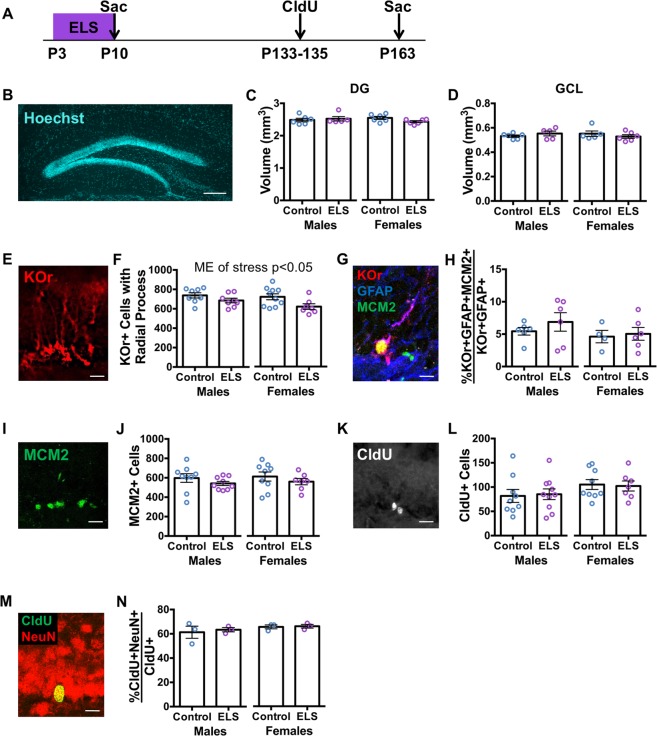
Early life stress predisposes to mental illness and behavioral dysfunction in adulthood, but the mechanisms underlying these persistent effects are poorly understood. Stress throughout life impairs the structure and function of the hippocampus, a brain system undergoing considerable development in early life. The long-term behavioral consequences of early life stress may therefore be due in part to interference with hippocampal development, in particular with assembly of the dentate gyrus (DG) region of the hippocampus. We investigated how early life stress produces long-term alterations in DG structure by examining DG assembly and the generation of a stable adult stem cell pool in routine housing and after stress induced by the limited bedding/nesting paradigm in mice. We found that early life stress leads to a more immature, proliferative DG than would be expected for the animal's age immediately after stress exposure, suggesting that early life stress delays DG development. Adult animals exposed to early life stress exhibited a reduction in the number of DG stem cells, but unchanged neurogenesis suggesting a depletion of the stem cell pool with compensation in the birth and survival of adult-born neurons. These results suggest a developmental mechanism by which early life stress can induce long-term changes in hippocampal function by interfering with DG assembly and ultimately diminishing the adult stem cell pool.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
The postnatal origin of adult neural stem cells and the effects of glucocorticoids on their genesis.Behav Brain Res. 2015 Feb 15;279:166-76. doi: 10.1016/j.bbr.2014.11.013. Epub 2014 Nov 12. Behav Brain Res. 2015. PMID: 25446750
-
Ablation of proliferating neural stem cells during early life is sufficient to reduce adult hippocampal neurogenesis.Hippocampus. 2018 Aug;28(8):586-601. doi: 10.1002/hipo.22962. Hippocampus. 2018. PMID: 29742815 Free PMC article.
-
Chronic early life stress alters developmental and adult neurogenesis and impairs cognitive function in mice.Hippocampus. 2015 Mar;25(3):309-28. doi: 10.1002/hipo.22374. Epub 2014 Oct 30. Hippocampus. 2015. PMID: 25269685
-
Stress and hippocampal neurogenesis.Biol Psychiatry. 1999 Dec 1;46(11):1472-9. doi: 10.1016/s0006-3223(99)00247-4. Biol Psychiatry. 1999. PMID: 10599477 Review.
-
Neural stem cell of the hippocampus: development, physiology regulation, and dysfunction in disease.Curr Top Dev Biol. 2014;107:183-206. doi: 10.1016/B978-0-12-416022-4.00007-X. Curr Top Dev Biol. 2014. PMID: 24439807 Review.
Cited by
-
Current Understanding of the Roles of Gut-Brain Axis in the Cognitive Deficits Caused by Perinatal Stress Exposure.Cells. 2023 Jun 28;12(13):1735. doi: 10.3390/cells12131735. Cells. 2023. PMID: 37443769 Free PMC article. Review.
-
Glucocorticoid effects on the brain: from adaptive developmental plasticity to allostatic overload.J Exp Biol. 2024 Mar 7;227(Suppl_1):jeb246128. doi: 10.1242/jeb.246128. Epub 2024 Mar 7. J Exp Biol. 2024. PMID: 38449327 Free PMC article. Review.
-
Olfactory neurogenesis plays different parts at successive stages of life, implications for mental health.Front Neural Circuits. 2024 Aug 8;18:1467203. doi: 10.3389/fncir.2024.1467203. eCollection 2024. Front Neural Circuits. 2024. PMID: 39175668 Free PMC article. Review.
-
Early Life Stress Induces Different Behaviors in Adolescence and Adulthood May Related With Abnormal Medial Prefrontal Cortex Excitation/Inhibition Balance.Front Neurosci. 2022 Jan 4;15:720286. doi: 10.3389/fnins.2021.720286. eCollection 2021. Front Neurosci. 2022. PMID: 35058738 Free PMC article.
-
Insulin and obesity transform hypothalamic-pituitary-adrenal axis stemness and function in a hyperactive state.Mol Metab. 2021 Jan;43:101112. doi: 10.1016/j.molmet.2020.101112. Epub 2020 Nov 4. Mol Metab. 2021. PMID: 33157254 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
