

Molecular Studies of the Protein Complexes Involving Cis-Prenyltransferase in Guayule (Parthenium argentatum), an Alternative Rubber-Producing Plant
- PMID: 30858856
- PMCID: PMC6397875
- DOI: 10.3389/fpls.2019.00165
Molecular Studies of the Protein Complexes Involving Cis-Prenyltransferase in Guayule (Parthenium argentatum), an Alternative Rubber-Producing Plant
Abstract
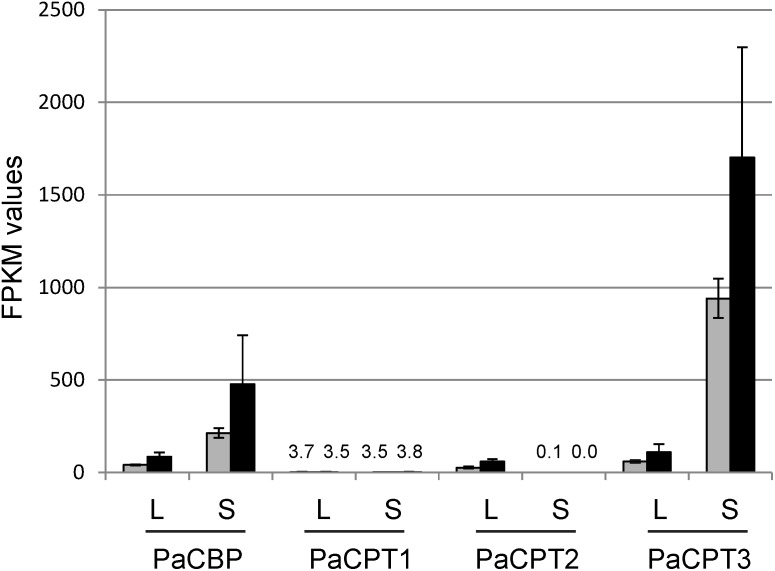
Guayule (Parthenium argentatum) is a perennial shrub in the Asteraceae family and synthesizes a high quality, hypoallergenic cis-1,4-polyisoprene (or natural rubber; NR). Despite its potential to be an alternative NR supplier, the enzymes for cis-polyisoprene biosynthesis have not been comprehensively studied in guayule. Recently, implications of the protein complex involving cis-prenyltransferases (CPTs) and CPT-Binding Proteins (CBPs) in NR biosynthesis were shown in lettuce and dandelion, but such protein complexes have yet to be examined in guayule. Here, we identified four guayule genes - three PaCPTs (PaCPT1-3) and one PaCBP, whose protein products organize PaCPT/PaCBP complexes. Co-expression of both PaCBP and each of the PaCPTs could complemented the dolichol (a short cis-polyisoprene)-deficient yeast, whereas the individual expressions could not. Microsomes from the PaCPT/PaCBP-expressing yeast efficiently incorporated 14C-isopentenyl diphosphate into dehydrodolichyl diphosphates; however, NR with high molecular weight could not be synthesized in in vitro assays. Furthermore, co-immunoprecipitation and split-ubiquitin yeast 2-hybrid assays using PaCPTs and PaCBP confirmed the formation of protein complexes. Of the three PaCPTs, guayule transcriptomics analysis indicated that the PaCPT3 is predominantly expressed in stem and induced by cold-stress, suggesting its involvement in NR biosynthesis. The comprehensive analyses of these PaCPTs and PaCBP here provide the foundational knowledge to generate a high NR-yielding guayule.
Keywords: cis-prenyltransferase; guayule (Parthenium argentatum); polyisoprene; protein complex; terpenoid.
Figures





Similar articles
-
Natural rubber biosynthesis in plants, the rubber transferase complex, and metabolic engineering progress and prospects.Plant Biotechnol J. 2019 Nov;17(11):2041-2061. doi: 10.1111/pbi.13181. Epub 2019 Jun 26. Plant Biotechnol J. 2019. PMID: 31150158 Free PMC article. Review.
-
Reconstitution of prenyltransferase activity on nanodiscs by components of the rubber synthesis machinery of the Para rubber tree and guayule.Sci Rep. 2022 Mar 8;12(1):3734. doi: 10.1038/s41598-022-07564-y. Sci Rep. 2022. PMID: 35260628 Free PMC article.
-
New insights into natural rubber biosynthesis from rubber-deficient lettuce mutants expressing goldenrod or guayule cis-prenyltransferase.New Phytol. 2023 Aug;239(3):1098-1111. doi: 10.1111/nph.18994. Epub 2023 May 29. New Phytol. 2023. PMID: 37247337
-
Elucidation of rubber biosynthesis and accumulation in the rubber producing shrub, guayule (Parthenium argentatum Gray).Planta. 2018 Feb;247(2):513-526. doi: 10.1007/s00425-017-2804-7. Epub 2017 Nov 7. Planta. 2018. PMID: 29116401
-
Guayule (Parthenium argentatum A. Gray), a Renewable Resource for Natural Polyisoprene and Resin: Composition, Processes and Applications.Molecules. 2021 Jan 27;26(3):664. doi: 10.3390/molecules26030664. Molecules. 2021. PMID: 33513965 Free PMC article. Review.
Cited by
-
Germacrene A Synthases for Sesquiterpene Lactone Biosynthesis Are Expressed in Vascular Parenchyma Cells Neighboring Laticifers in Lettuce.Plants (Basel). 2022 Apr 28;11(9):1192. doi: 10.3390/plants11091192. Plants (Basel). 2022. PMID: 35567193 Free PMC article.
-
Another level of complex-ity: The role of metabolic channeling and metabolons in plant terpenoid metabolism.Front Plant Sci. 2022 Aug 10;13:954083. doi: 10.3389/fpls.2022.954083. eCollection 2022. Front Plant Sci. 2022. PMID: 36035727 Free PMC article. Review.
-
Downregulation of Squalene Synthase Broadly Impacts Isoprenoid Biosynthesis in Guayule.Metabolites. 2022 Mar 29;12(4):303. doi: 10.3390/metabo12040303. Metabolites. 2022. PMID: 35448489 Free PMC article.
-
Natural rubber biosynthesis in plants, the rubber transferase complex, and metabolic engineering progress and prospects.Plant Biotechnol J. 2019 Nov;17(11):2041-2061. doi: 10.1111/pbi.13181. Epub 2019 Jun 26. Plant Biotechnol J. 2019. PMID: 31150158 Free PMC article. Review.
-
Structure-function studies of ultrahigh molecular weight isoprenes provide key insights into their biosynthesis.Commun Biol. 2021 Feb 16;4(1):215. doi: 10.1038/s42003-021-01739-5. Commun Biol. 2021. PMID: 33594248 Free PMC article.
References
-
- Asawatreratanakul K., Zhang Y. W., Wititsuwannakul D., Wititsuwannakul R., Takahashi S., Rattanapittayaporn A., et al. (2003). Molecular cloning, expression, and characterization of cDNA encoding cis-prenyltransferases from Hevea brasiliensis. Eur. J. Biochem. 270 4671–4680. 10.1046/j.1432-1033.2003.03863.x - DOI - PubMed
-
- Cantú D. J., Angulo-Sánchez J. L., Rodríguez-García R., Kuruvadi S. (1997). Seasonal growth, rubber and resin yield characteristics of guayule under natural environmental conditions. Ind. Crops Prod. 6 131–137. 10.1016/S0926-6690(96)00211-7 - DOI
LinkOut - more resources
Full Text Sources
Molecular Biology Databases