

The histone methyltransferase G9a regulates tolerance to oxidative stress-induced energy consumption
- PMID: 30860988
- PMCID: PMC6413895
- DOI: 10.1371/journal.pbio.2006146
The histone methyltransferase G9a regulates tolerance to oxidative stress-induced energy consumption
Abstract
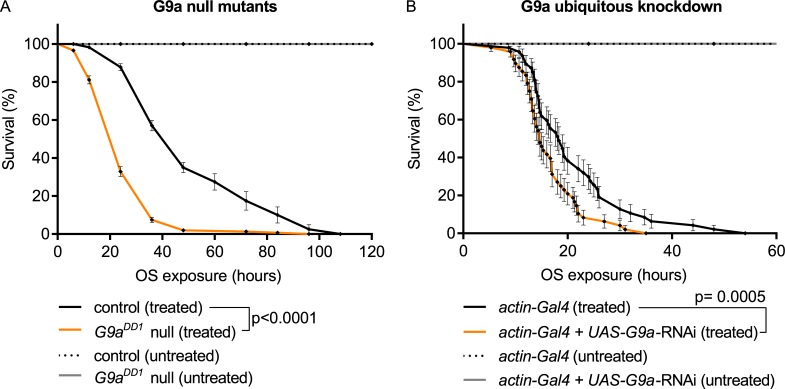
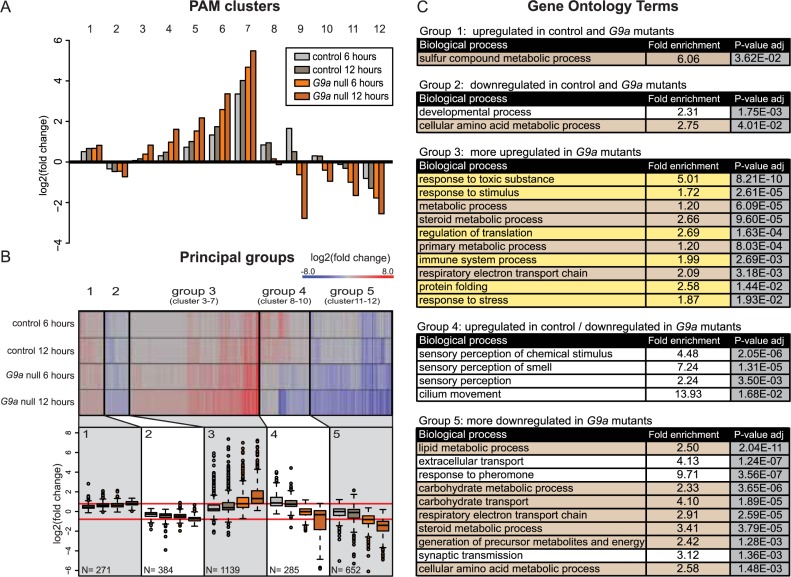
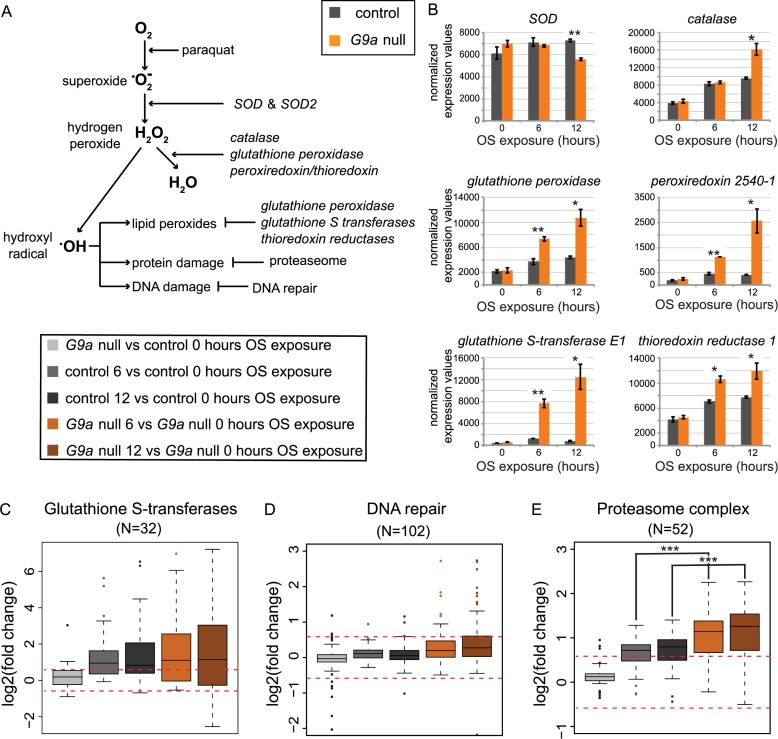
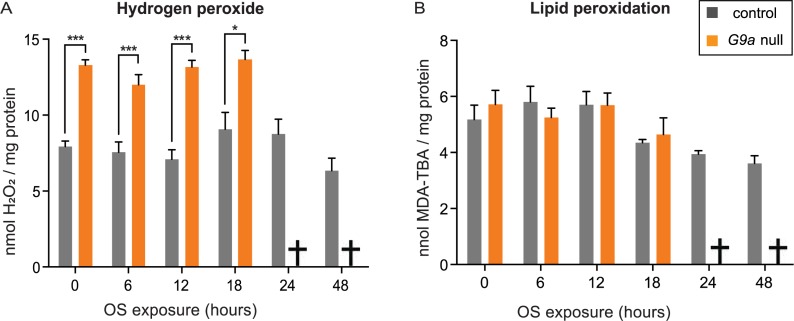
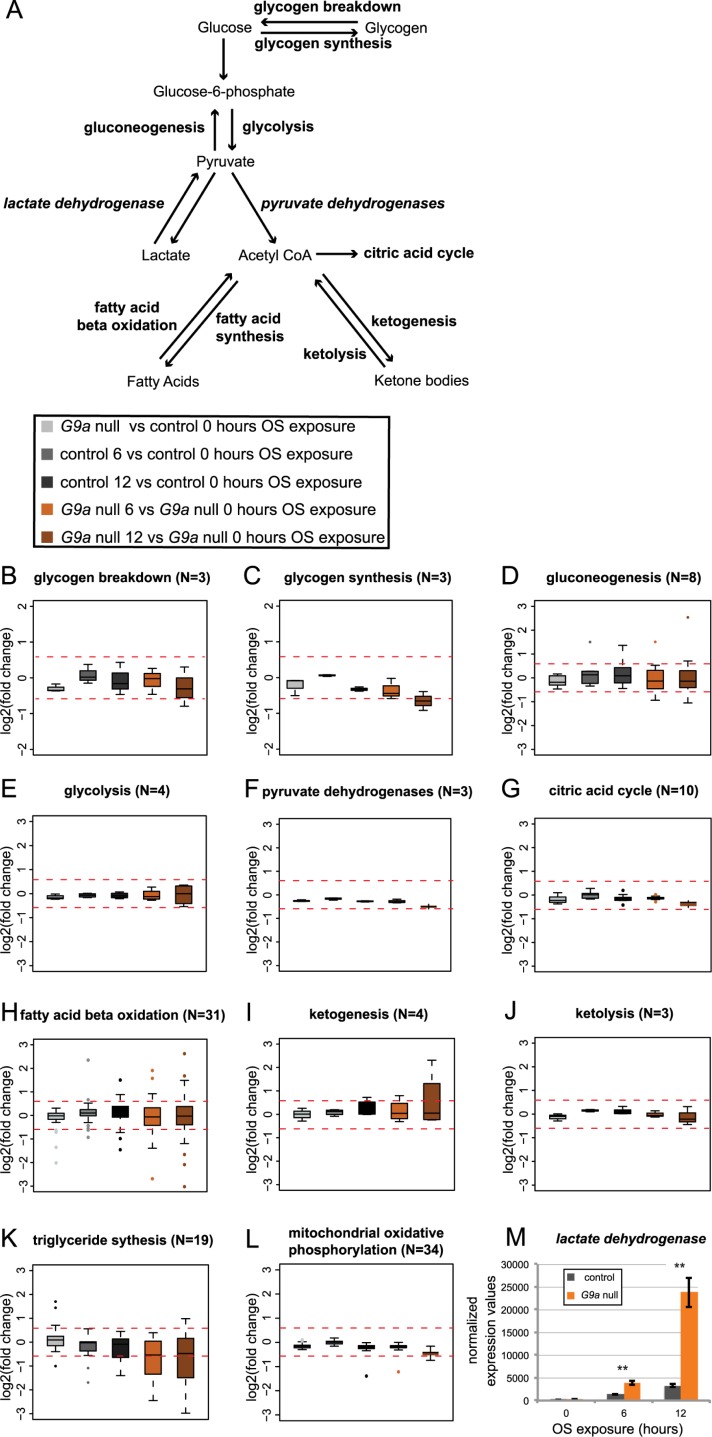
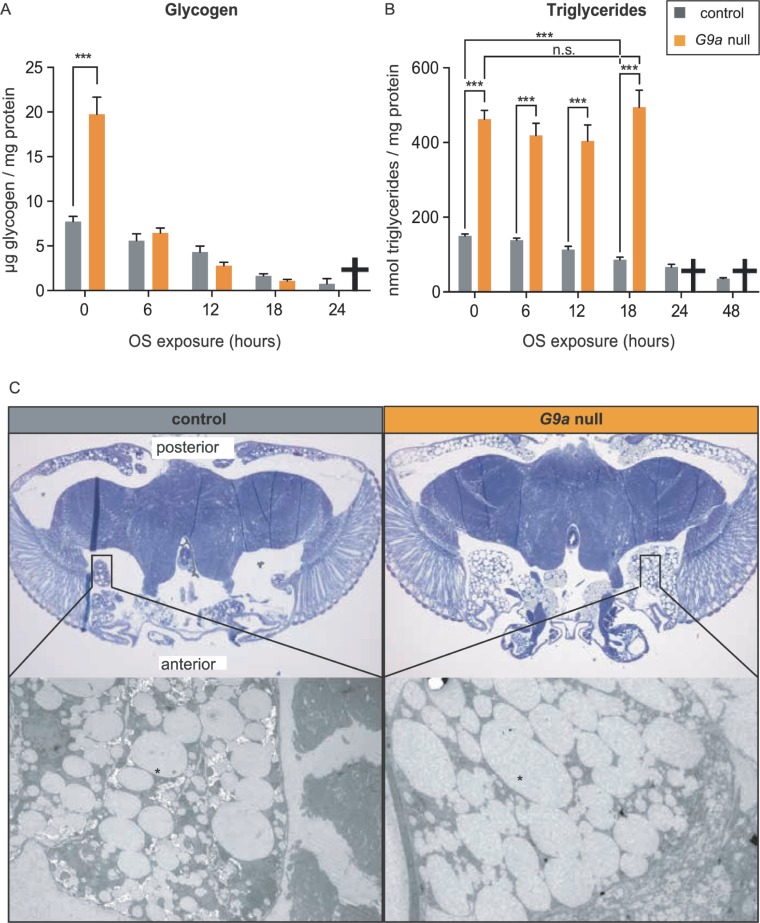
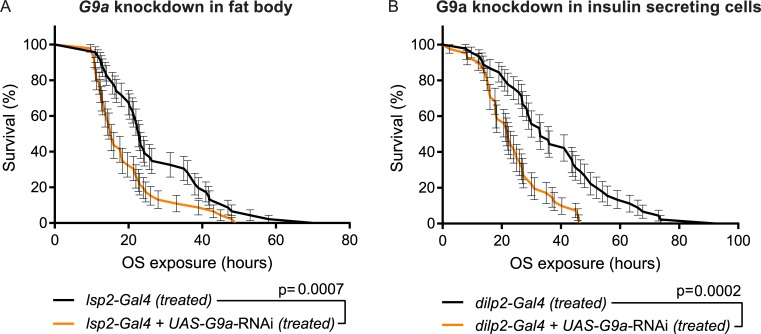
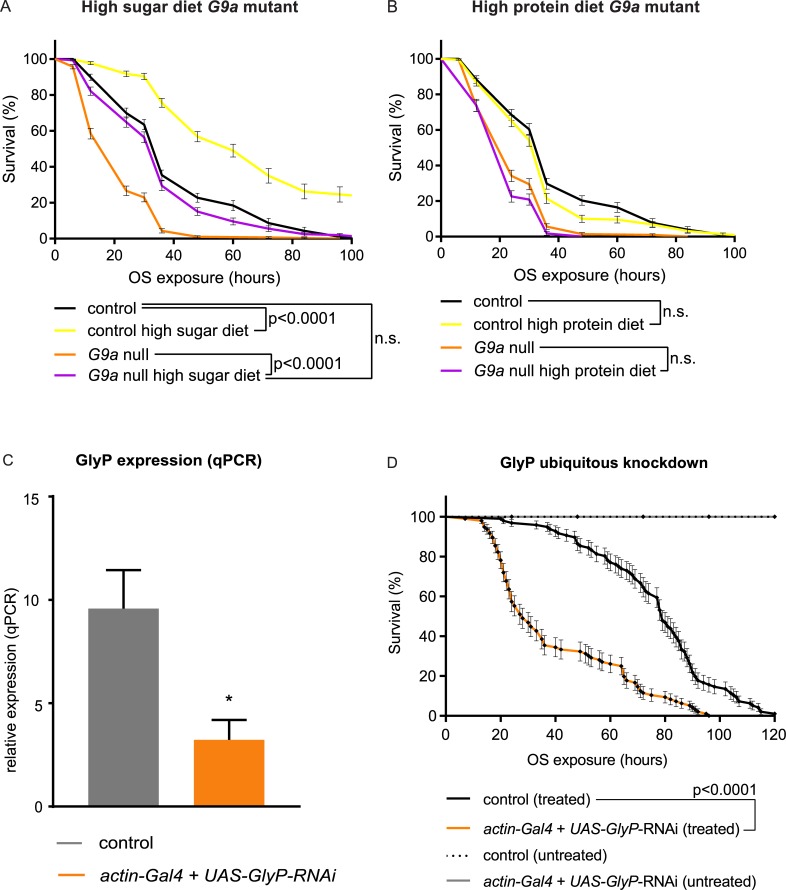
Stress responses are crucial processes that require activation of genetic programs that protect from the stressor. Stress responses are also energy consuming and can thus be deleterious to the organism. The mechanisms coordinating energy consumption during stress response in multicellular organisms are not well understood. Here, we show that loss of the epigenetic regulator G9a in Drosophila causes a shift in the transcriptional and metabolic responses to oxidative stress (OS) that leads to decreased survival time upon feeding the xenobiotic paraquat. During OS exposure, G9a mutants show overactivation of stress response genes, rapid depletion of glycogen, and inability to access lipid energy stores. The OS survival deficiency of G9a mutants can be rescued by a high-sugar diet. Control flies also show improved OS survival when fed a high-sugar diet, suggesting that energy availability is generally a limiting factor for OS tolerance. Directly limiting access to glycogen stores by knocking down glycogen phosphorylase recapitulates the OS-induced survival defects of G9a mutants. We propose that G9a mutants are sensitive to stress because they experience a net reduction in available energy due to (1) rapid glycogen use, (2) an inability to access lipid energy stores, and (3) an overinduced transcriptional response to stress that further exacerbates energy demands. This suggests that G9a acts as a critical regulatory hub between the transcriptional and metabolic responses to OS. Our findings, together with recent studies that established a role for G9a in hypoxia resistance in cancer cell lines, suggest that G9a is of wide importance in controlling the cellular and organismal response to multiple types of stress.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
Epigenetic regulator G9a provides glucose as a sweet key to stress resistance.PLoS Biol. 2019 Apr 19;17(4):e3000236. doi: 10.1371/journal.pbio.3000236. eCollection 2019 Apr. PLoS Biol. 2019. PMID: 31002662 Free PMC article.
References
-
- Kültz D. Molecular and evolutionary basis of the cellular stress response. Annu Rev Physiol. 2005;67(1):225–57. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
