

Epithelial (E)-Cadherin is a Novel Mediator of Platelet Aggregation and Clot Stability
- PMID: 30861547
- PMCID: PMC6599679
- DOI: 10.1055/s-0039-1679908
Epithelial (E)-Cadherin is a Novel Mediator of Platelet Aggregation and Clot Stability
Abstract
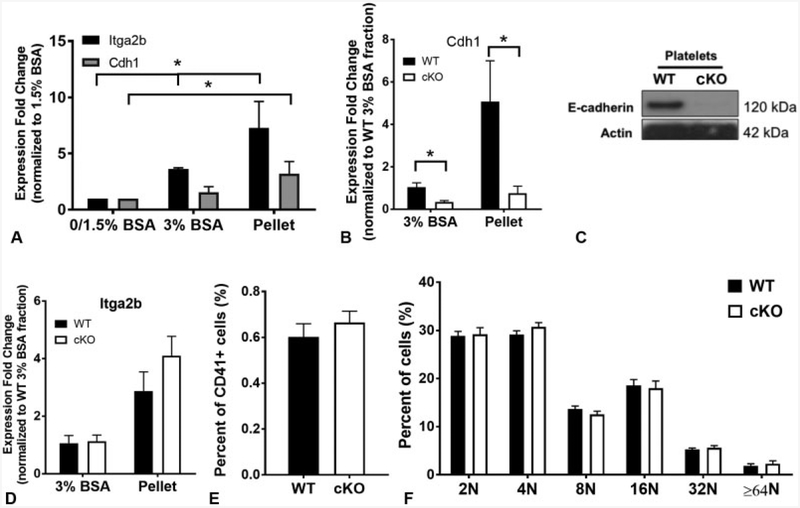
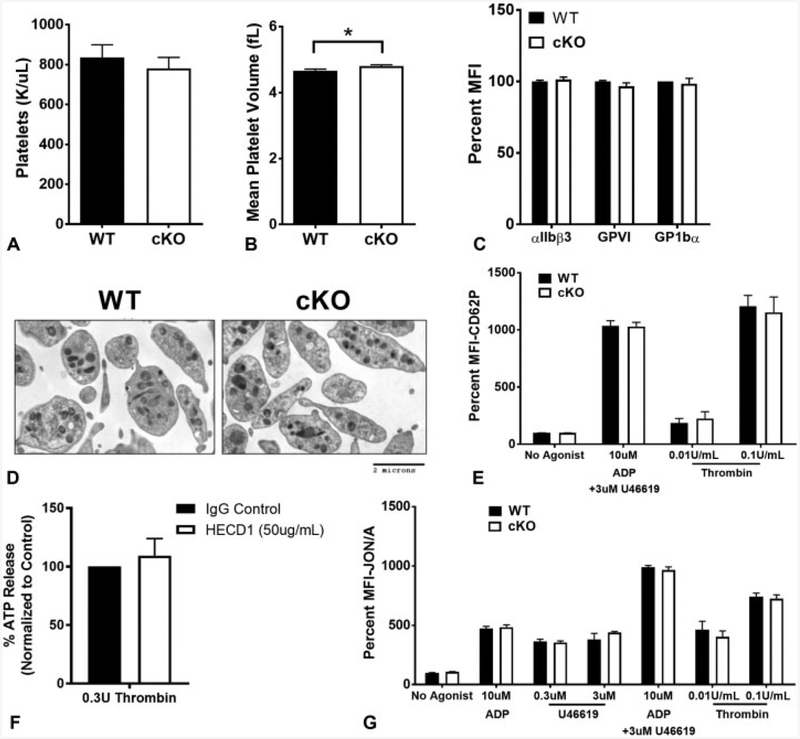
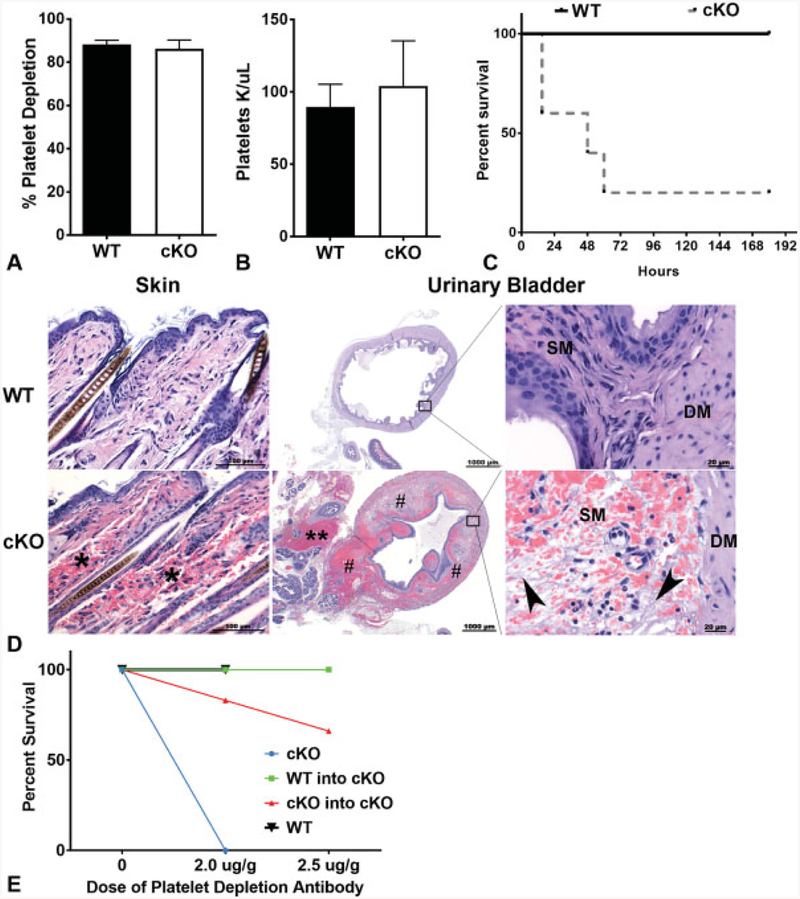
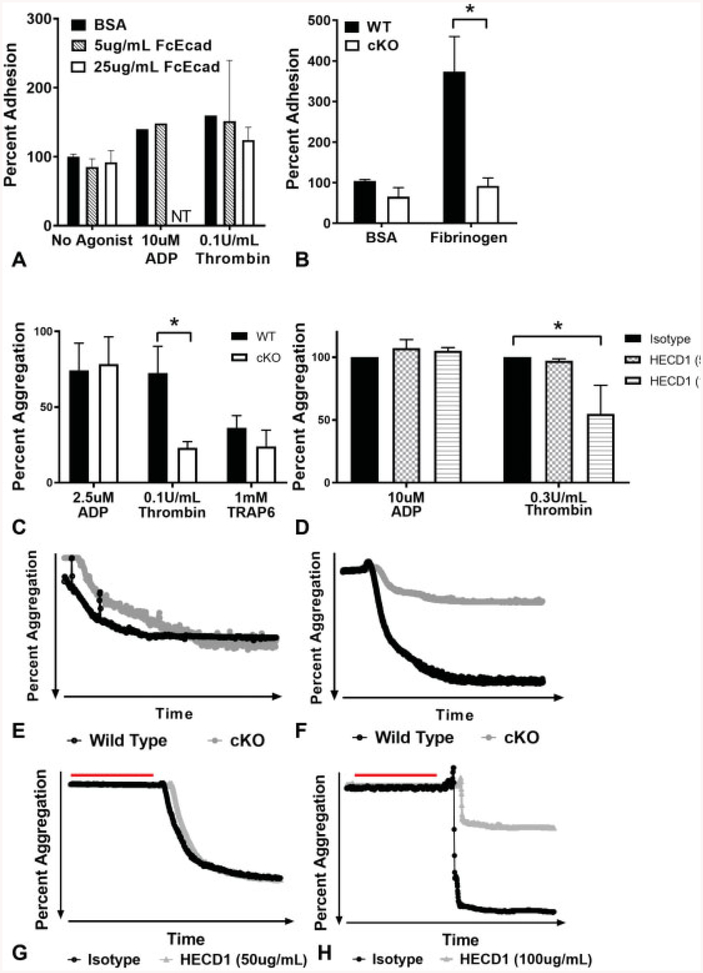
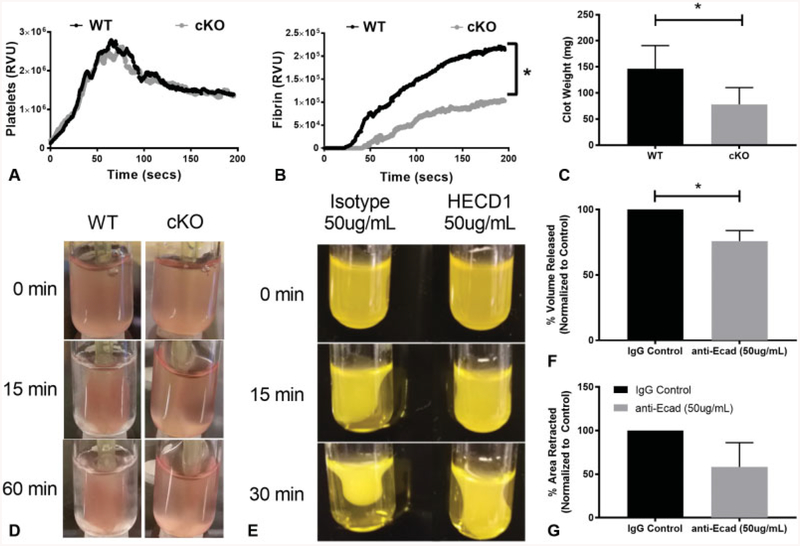
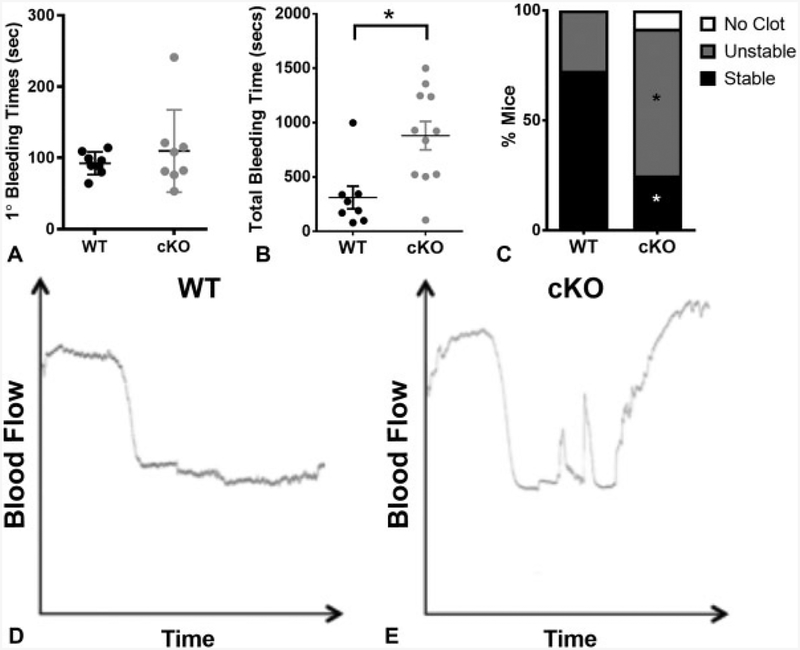
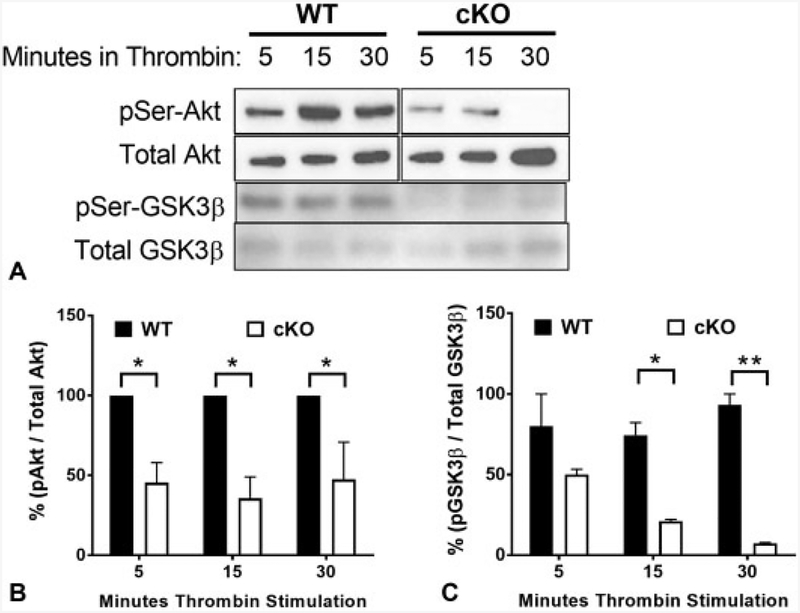
Cadherins play a major role in mediating cell-cell adhesion, which shares many parallels with platelet-platelet interactions during aggregate formation and clot stabilization. Platelets express epithelial (E)-cadherin, but its contribution to platelet function and/or platelet production is currently unknown. To assess the role of E-cadherin in platelet production and function in vitro and in vivo, we utilized a megakaryocyte-specific E-cadherin knockout mouse model. Loss of E-cadherin in megakaryocytes does not affect megakaryocyte maturation, platelet number or size. However, platelet dysfunction in the absence of E-cadherin is revealed when conditional knockout mice are challenged with acute antibody-mediated platelet depletion. Unlike wild-type mice that recover fully, knockout mice die within 72 hours post-antibody administration, likely from haemorrhage. Furthermore, conditional knockout mice have prolonged tail bleeding times, unstable clot formation, reduced clot retraction and reduced fibrin deposition in in vivo injury models. Murine platelet aggregation in vitro in response to thrombin and thrombin receptor activating peptide is compromised in E-cadherin null platelets, while aggregation in response to adenosine diphosphate (ADP) is not significantly different. Consistent with this, in vitro aggregation of primary human platelets in response to thrombin is decreased by an inhibitory E-cadherin antibody. Integrin activation and granule secretion in response to ADP and thrombin are not affected in E-cadherin null platelets, but Akt and glycogen synthase kinase 3β (GSK3β) activation are attenuated, suggesting a that E-cadherin contributes to aggregation, clot stabilization and retraction that is mediated by phosphoinositide 3-kinase/Akt/GSK3β signalling. In summary, E-cadherin plays a salient role in platelet aggregation and clot stability.
Georg Thieme Verlag KG Stuttgart · New York.
Conflict of interest statement
None declared.
Figures
References
-
- Signaling Du X. and regulation of the platelet glycoprotein Ib-IX-V complex. Curr Opin Hematol 2007;14(03):262–269 - PubMed
-
- Yang H, Reheman A, Chen P, et al. Fibrinogen and von Willebrand factor-independent platelet aggregation in vitro and in vivo. J Thromb Haemost 2006;4(10):2230–2237 - PubMed
-
- Nurden AT, Fiore M, Nurden P, Pillois X. Glanzmann thrombasthenia: a review of ITGA2B and ITGB3 defects with emphasis on variants, phenotypic variability, and mouse models. Blood 2011; 118(23):5996–6005 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
