

Highly Resolved Systems Biology to Dissect the Etioplast-to-Chloroplast Transition in Tobacco Leaves
- PMID: 30862726
- PMCID: PMC6501100
- DOI: 10.1104/pp.18.01432
Highly Resolved Systems Biology to Dissect the Etioplast-to-Chloroplast Transition in Tobacco Leaves
Abstract
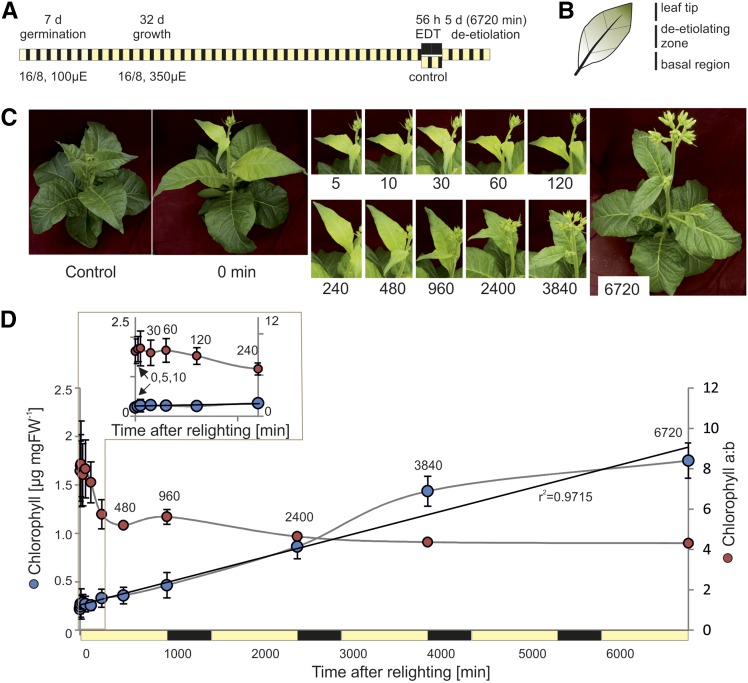
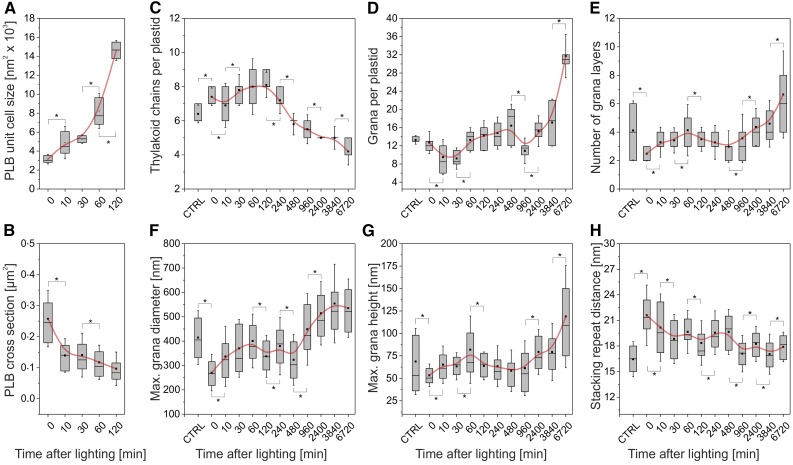
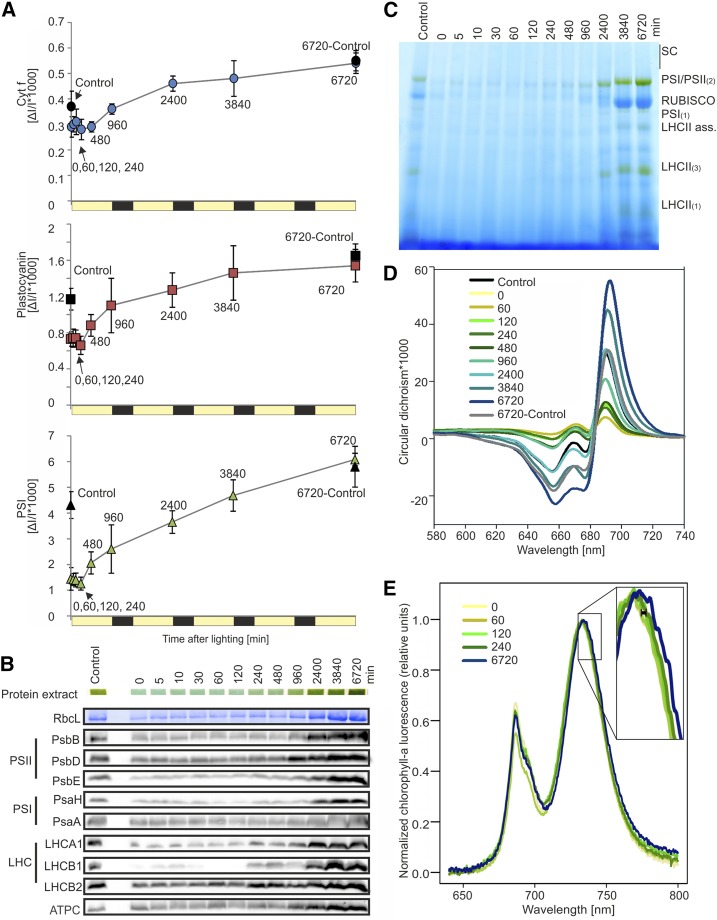
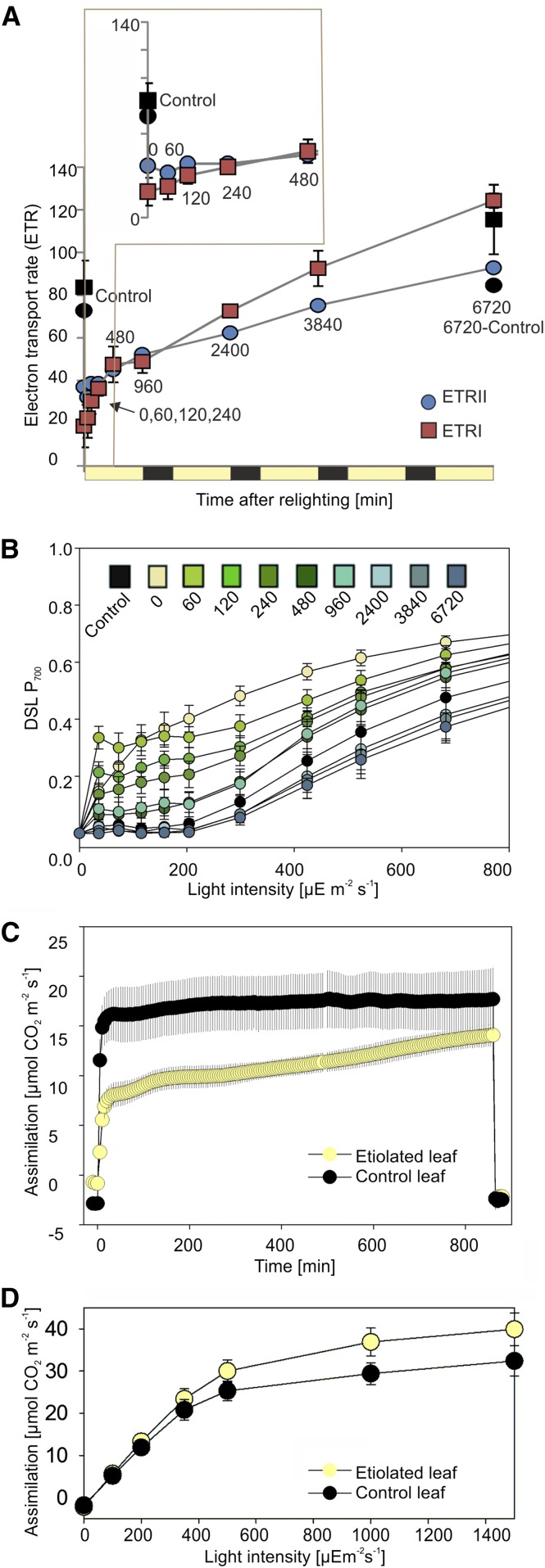
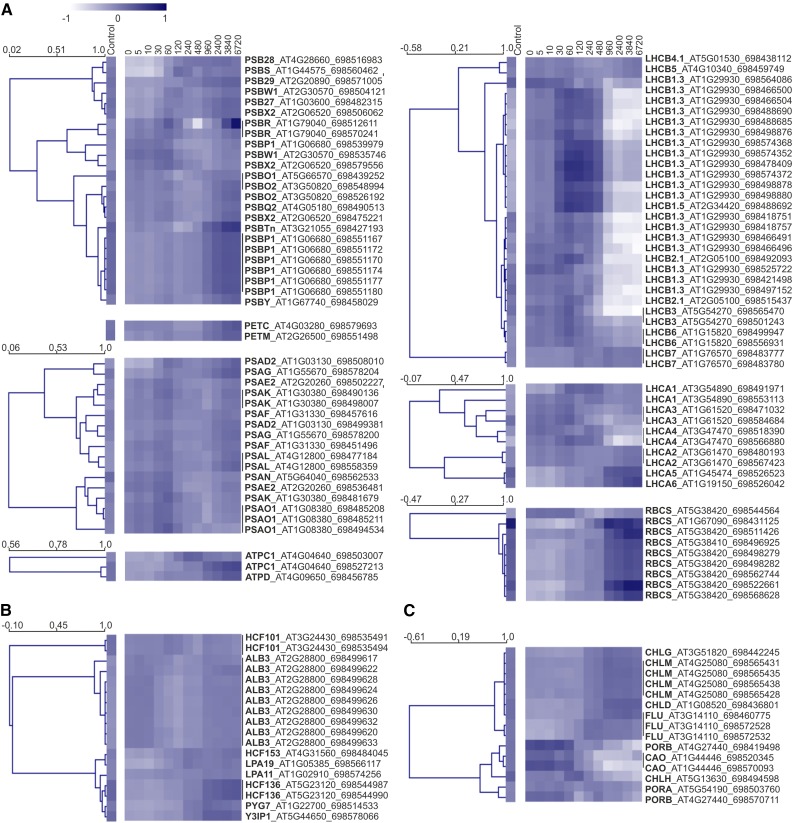
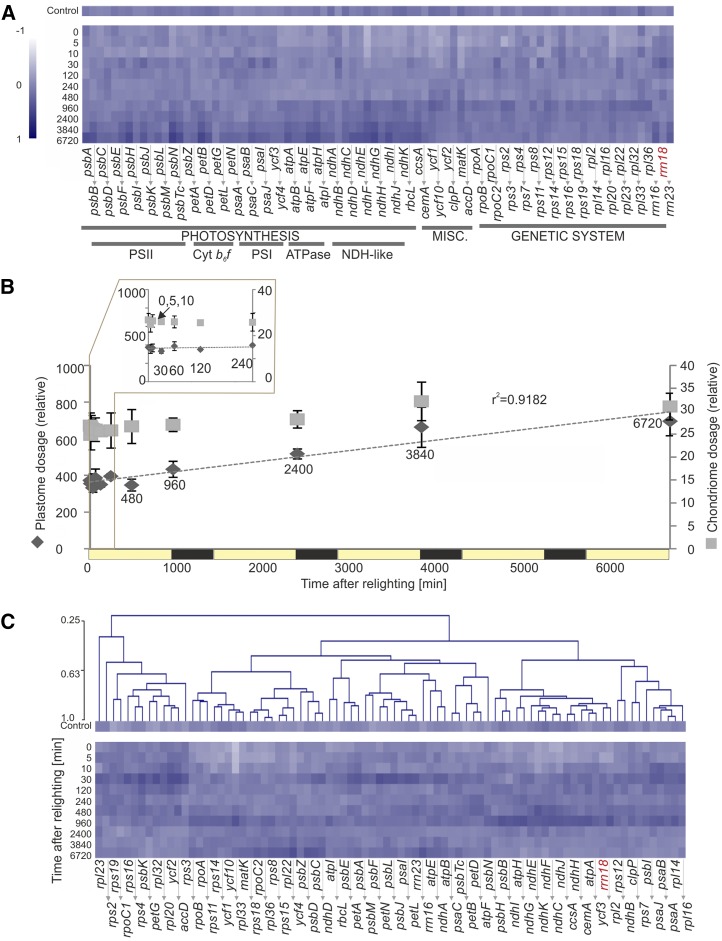
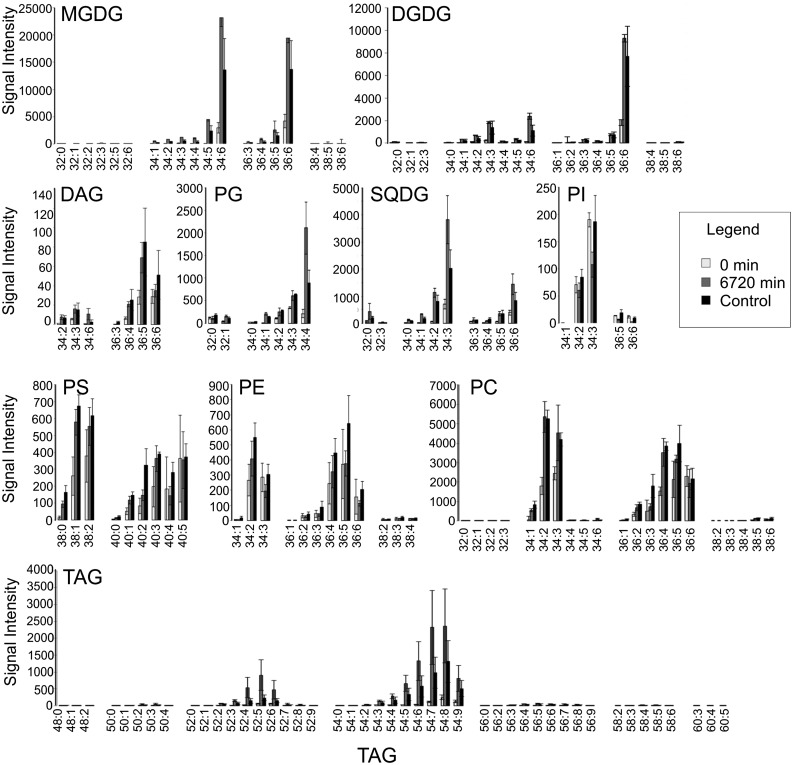
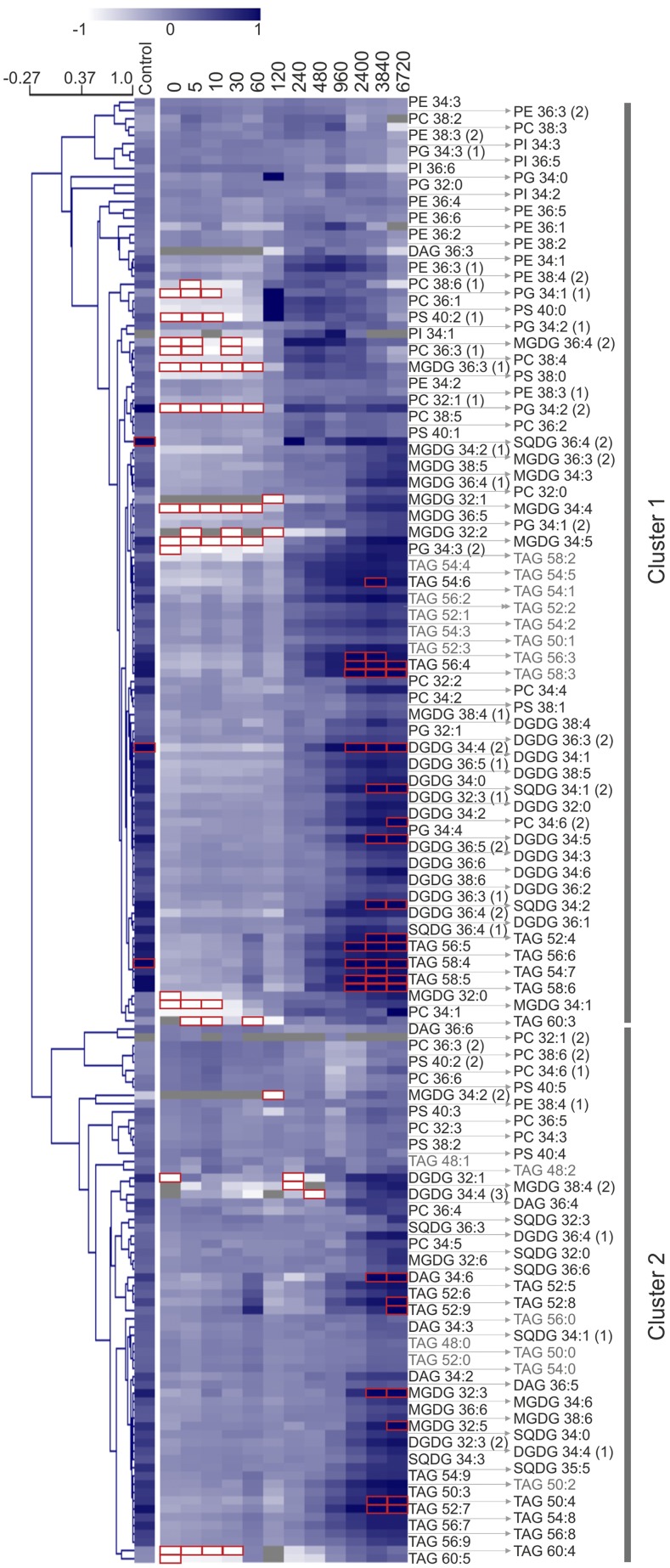
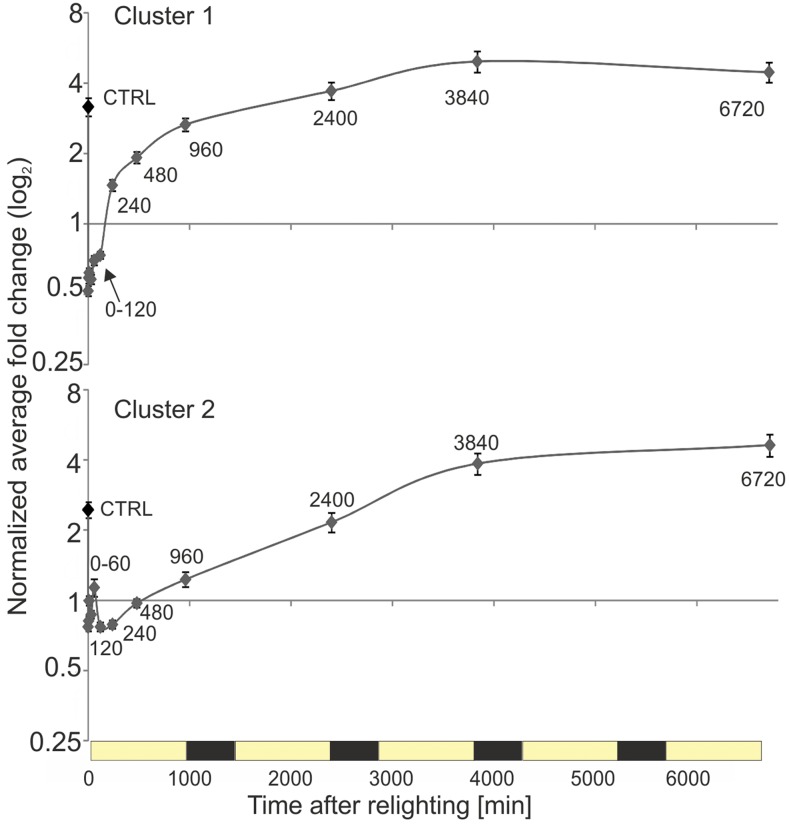
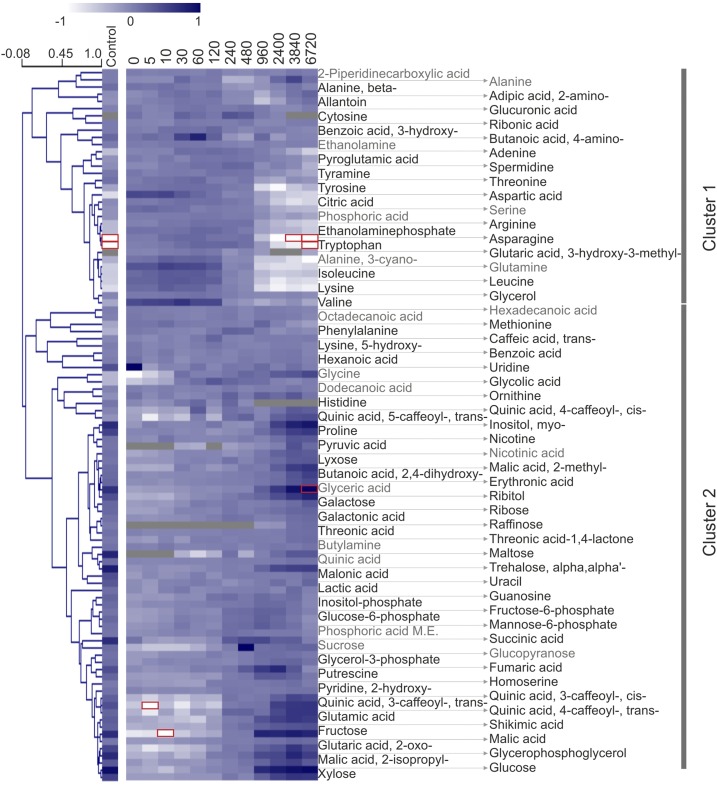
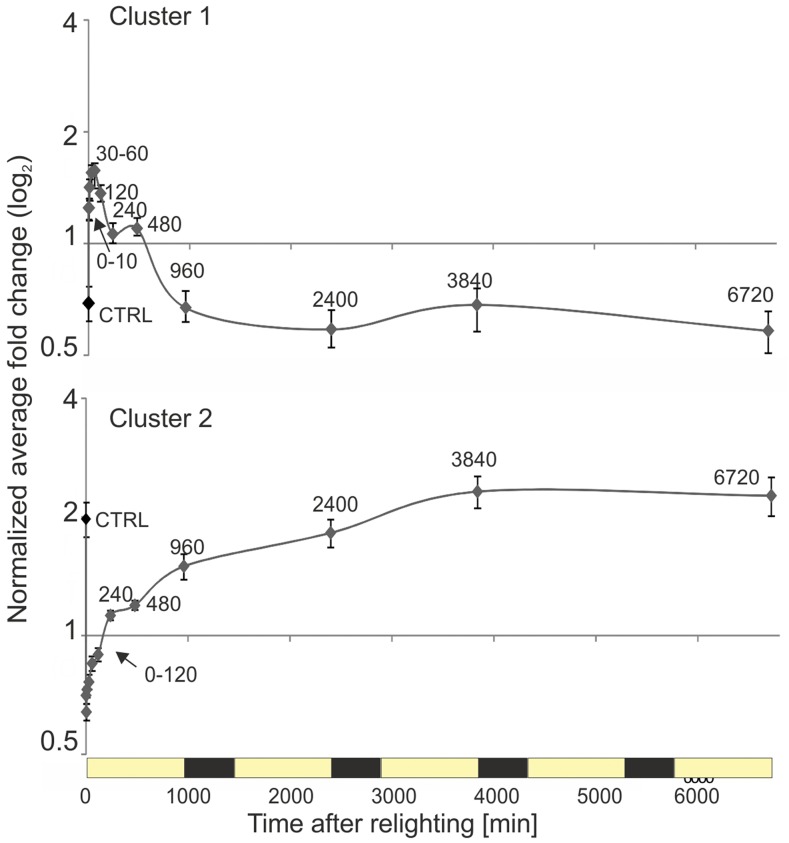
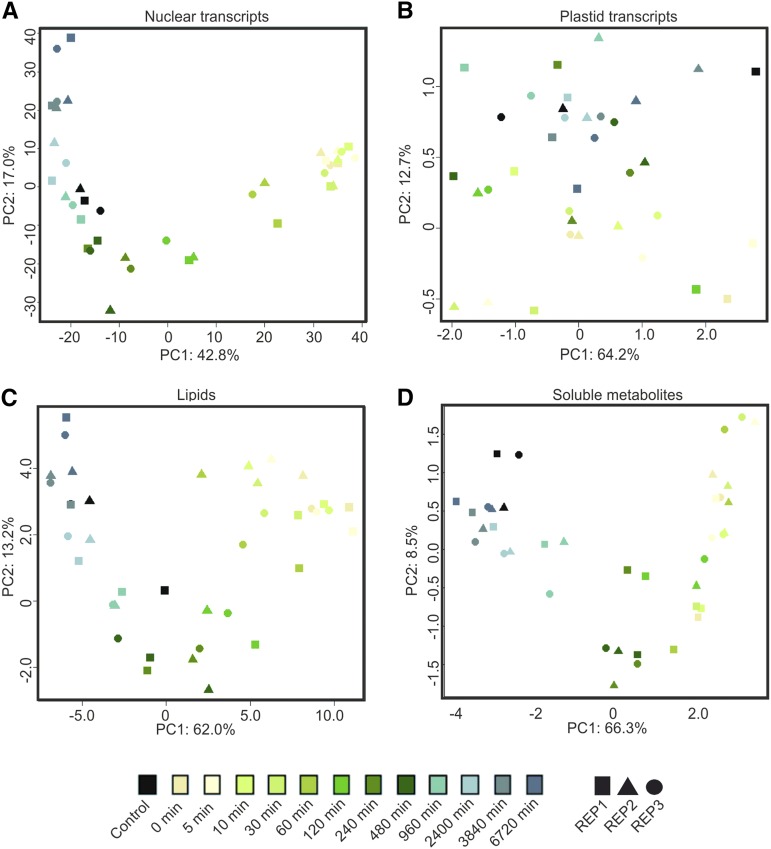
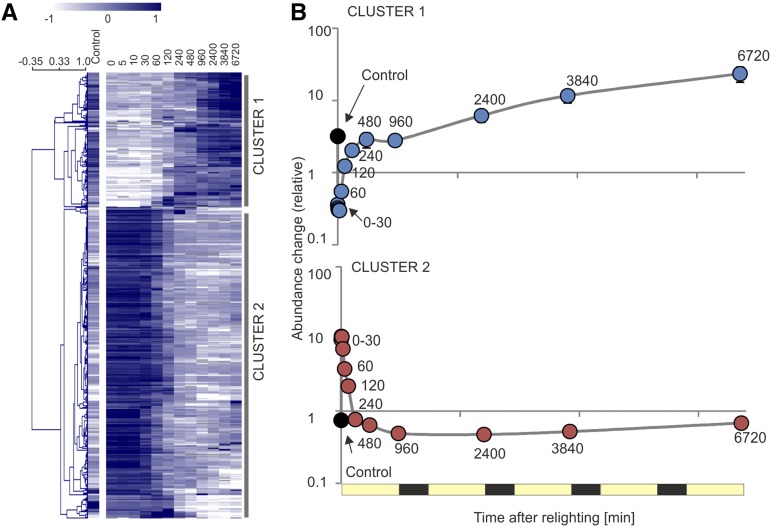
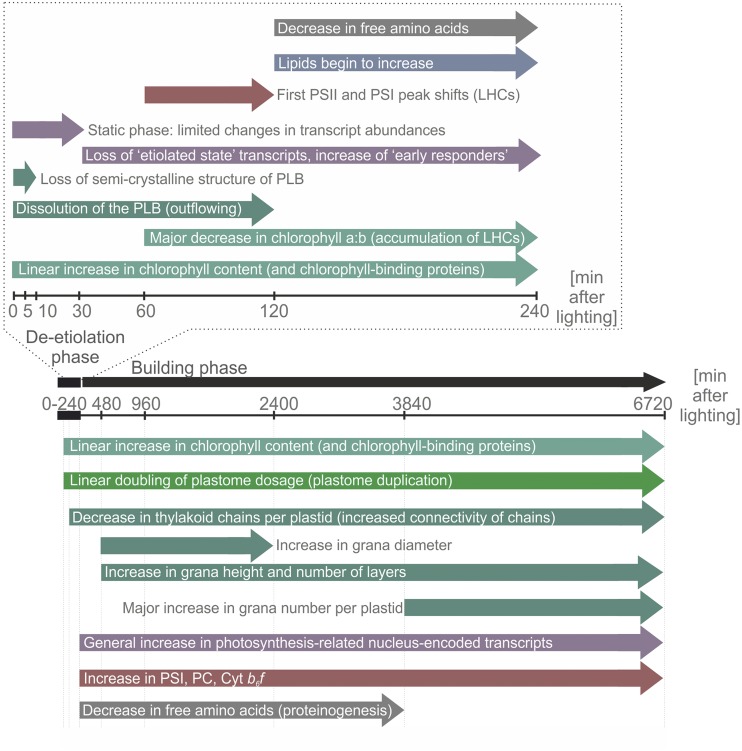
Upon exposure to light, plant cells quickly acquire photosynthetic competence by converting pale etioplasts into green chloroplasts. This developmental transition involves the de novo biogenesis of the thylakoid system and requires reprogramming of metabolism and gene expression. Etioplast-to-chloroplast differentiation involves massive changes in plastid ultrastructure, but how these changes are connected to specific changes in physiology, metabolism, and expression of the plastid and nuclear genomes is poorly understood. Here, we describe a new experimental system in the dicotyledonous model plant tobacco (Nicotiana tabacum) that allows us to study the leaf deetiolation process at the systems level. We have determined the accumulation kinetics of photosynthetic complexes, pigments, lipids, and soluble metabolites and recorded the dynamic changes in plastid ultrastructure and in the nuclear and plastid transcriptomes. Our data describe the greening process at high temporal resolution, resolve distinct genetic and metabolic phases during deetiolation, and reveal numerous candidate genes that may be involved in light-induced chloroplast development and thylakoid biogenesis.
© 2019 American Society of Plant Biologists. All Rights Reserved.
Figures

References
-
- Abramoff MD, Magalhães PJ, Ram SJ (2004) Image processing with ImageJ. Biophoton Int 11: 36–42
-
- Adam Z, Charuvi D, Tsabari O, Knopf RR, Reich Z (2011) Biogenesis of thylakoid networks in angiosperms: Knowns and unknowns. Plant Mol Biol 76: 221–234 - PubMed
-
- Aguettaz P, Seyer P, Pesey H, Lescure AM (1987) Relations between the plastid gene dosage and the levels of 16S rRNA and rbcL gene transcripts during amyloplast to chloroplast change in mixotrophic spinach cell suspensions. Plant Mol Biol 8: 169–177 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
