

Phosphorylated and aggregated TDP-43 with seeding properties are induced upon mutant Huntingtin (mHtt) polyglutamine expression in human cellular models
- PMID: 30863908
- PMCID: PMC11105362
- DOI: 10.1007/s00018-019-03059-8
Phosphorylated and aggregated TDP-43 with seeding properties are induced upon mutant Huntingtin (mHtt) polyglutamine expression in human cellular models
Abstract
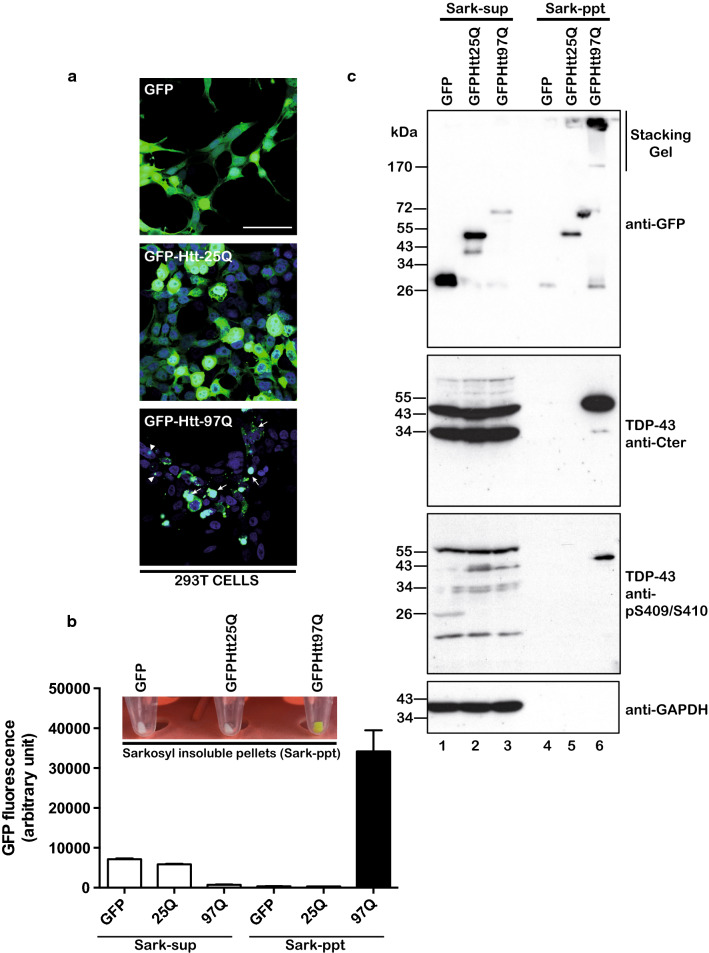
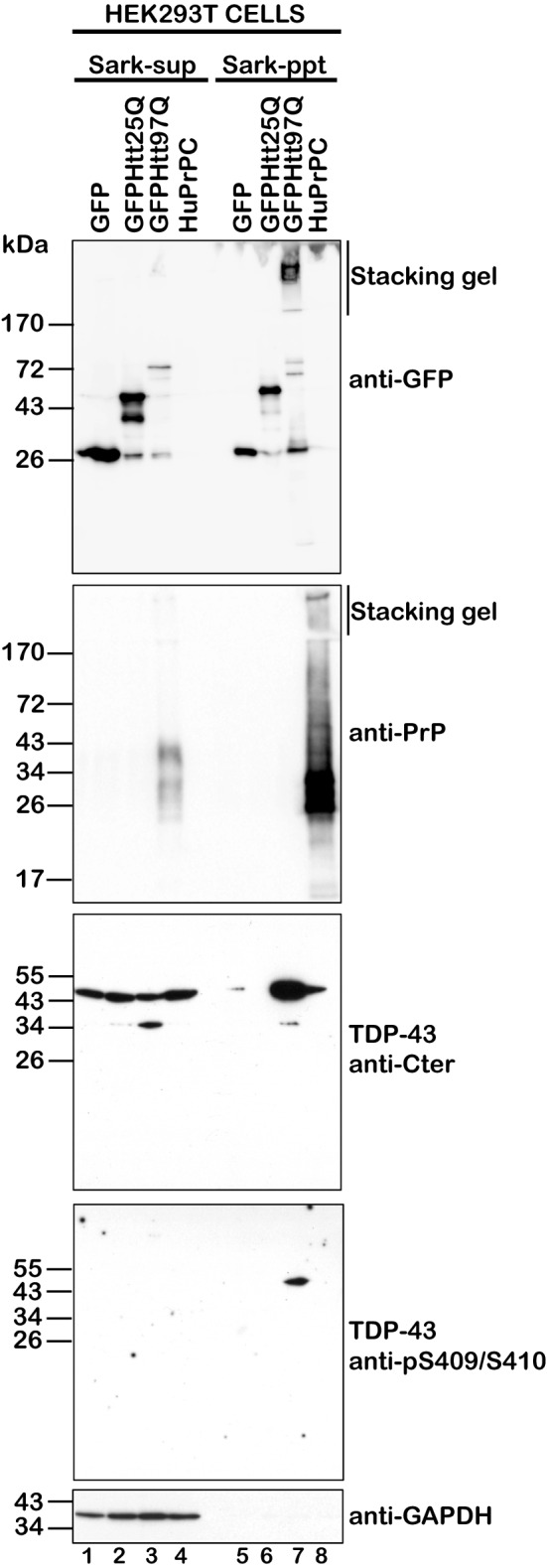
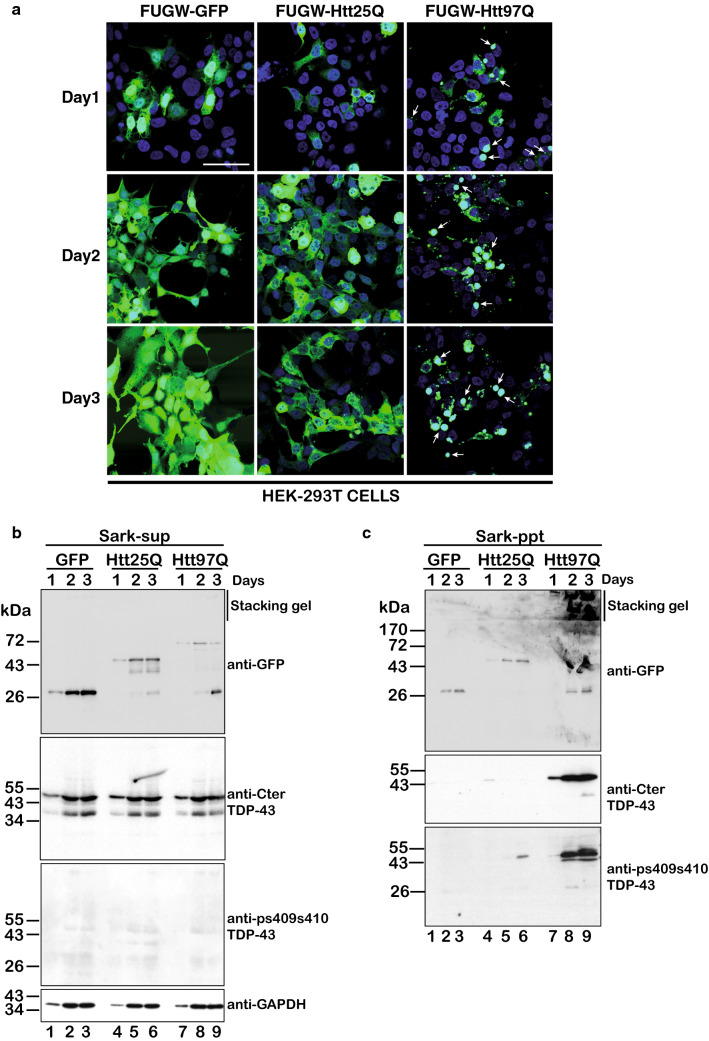
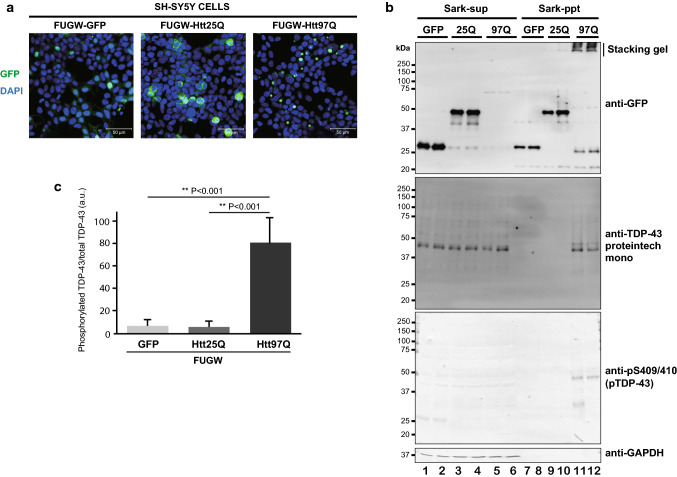
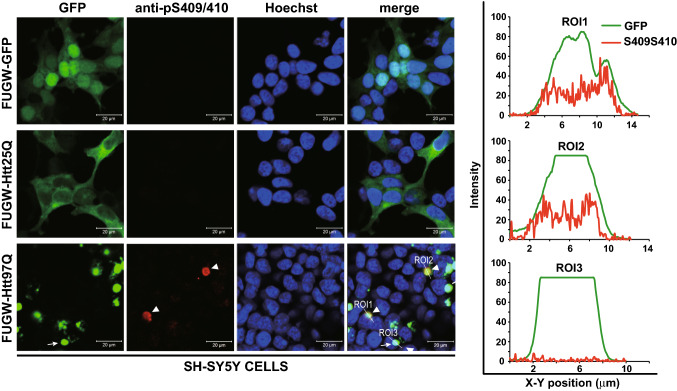
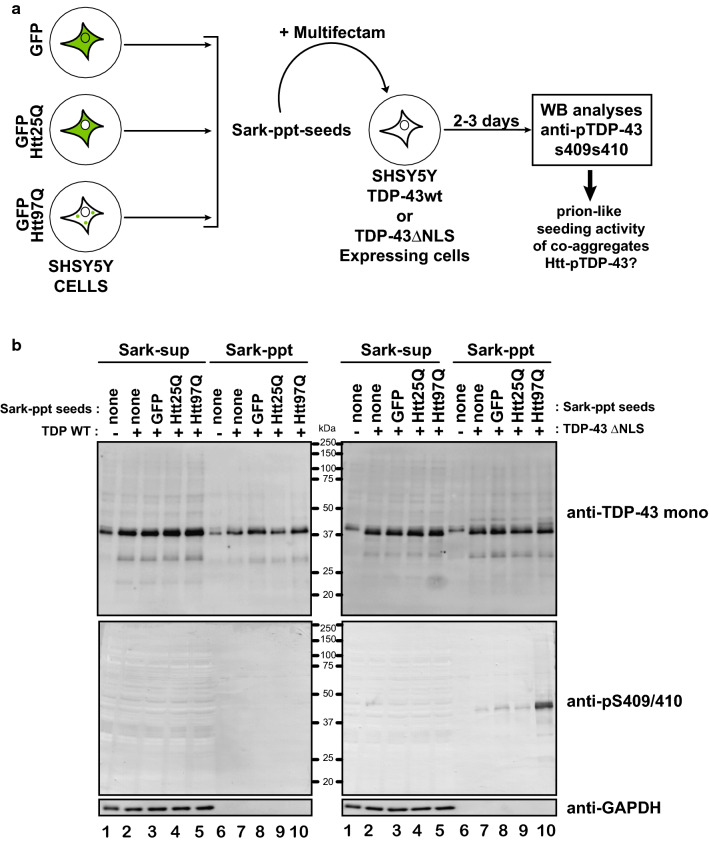
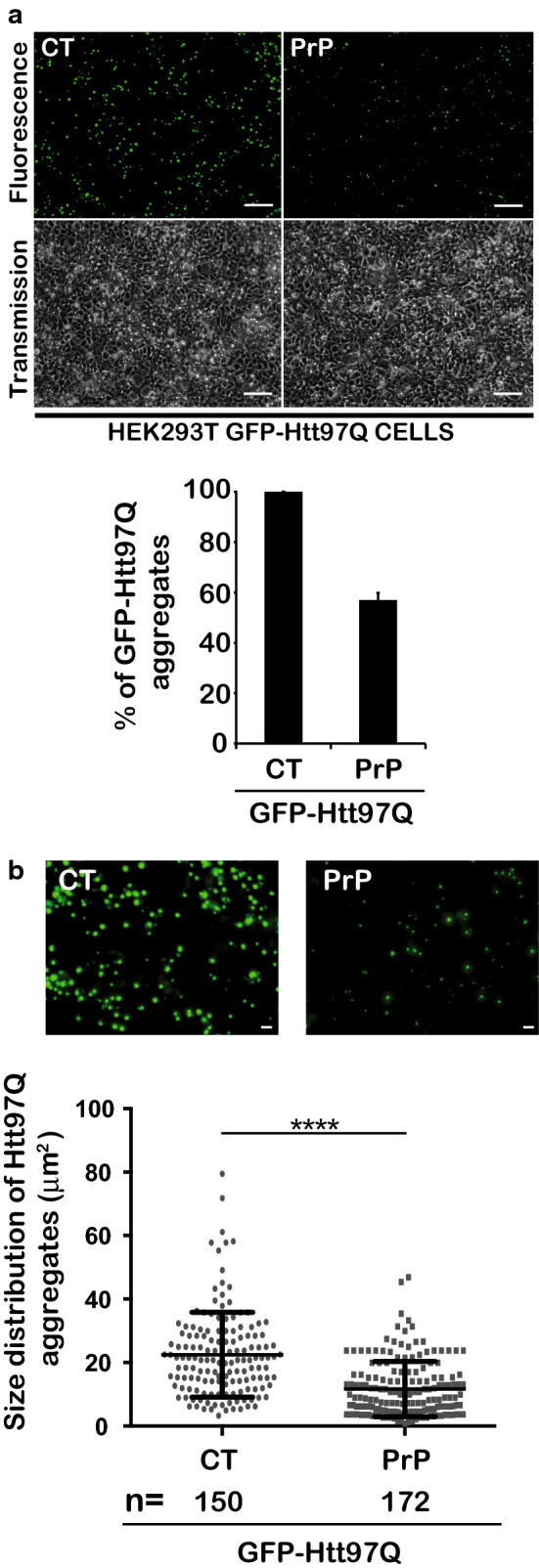
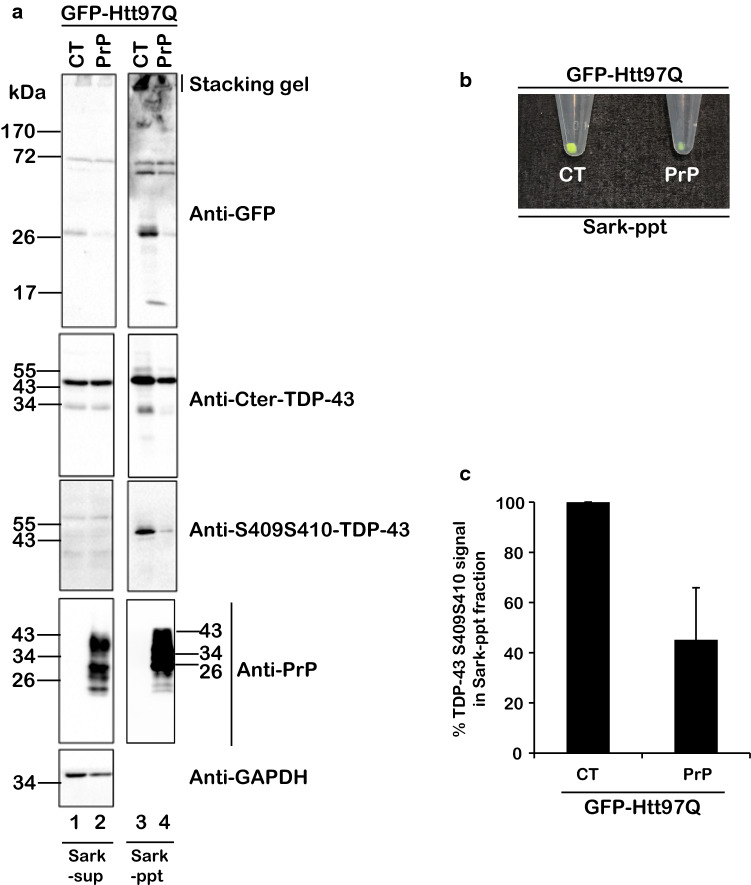
The Tar DNA-Binding Protein 43 (TDP-43) and its phosphorylated isoform (pTDP-43) are the major components associated with ubiquitin positive/Tau-negative inclusions found in neurons and glial cells of patients suffering of amyotrophic lateral sclerosis (ALS) or frontotemporal lobar degeneration-TDP-43 (FTLD-TDP). Many studies have revealed that TDP-43 is also in the protein inclusions associated with neurodegenerative conditions other than ALS and FTLD-TDP, thus suggesting that this protein may be involved in the pathogenesis of a variety of neurological disorders. In brains of Huntington-affected patients, pTDP-43 aggregates were shown to co-localize with mutant Huntingtin (mHtt) inclusions. Here, we show that expression of mHtt carrying 80-97 polyglutamines repeats in human cell cultures induces the aggregation and the phosphorylation of endogenous TDP-43, whereas non-pathological Htt with 25 polyglutamines repeats has no effect. Mutant Htt aggregation precedes accumulation of pTDP-43 and pTDP-43 co-localizes with mHtt inclusions reminding what it was previously described in brains of Huntington-affected patients. Detergent-insoluble fractions from cells expressing mHtt and containing mHtt-pTDP-43 co-aggregates can function as seeds for further TDP-43 aggregation in human cell culture. The human cellular prion protein PrPC was previously identified as a negative modulator of mHtt aggregation; here, we show that PrPC-mediated reduction of mHtt aggregation is tightly correlated with a decrease of TDP-43 aggregation and phosphorylation, thus confirming the close relationships between TDP-43 and mHtt.
Keywords: ALS; Huntingtin; Huntington; Phosphorylation; Prion; Seeds; TDP-43.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
