

Cross-reactive antibodies binding to H4 hemagglutinin protect against a lethal H4N6 influenza virus challenge in the mouse model
- PMID: 30866770
- PMCID: PMC6455122
- DOI: 10.1080/22221751.2018.1564369
Cross-reactive antibodies binding to H4 hemagglutinin protect against a lethal H4N6 influenza virus challenge in the mouse model
Abstract
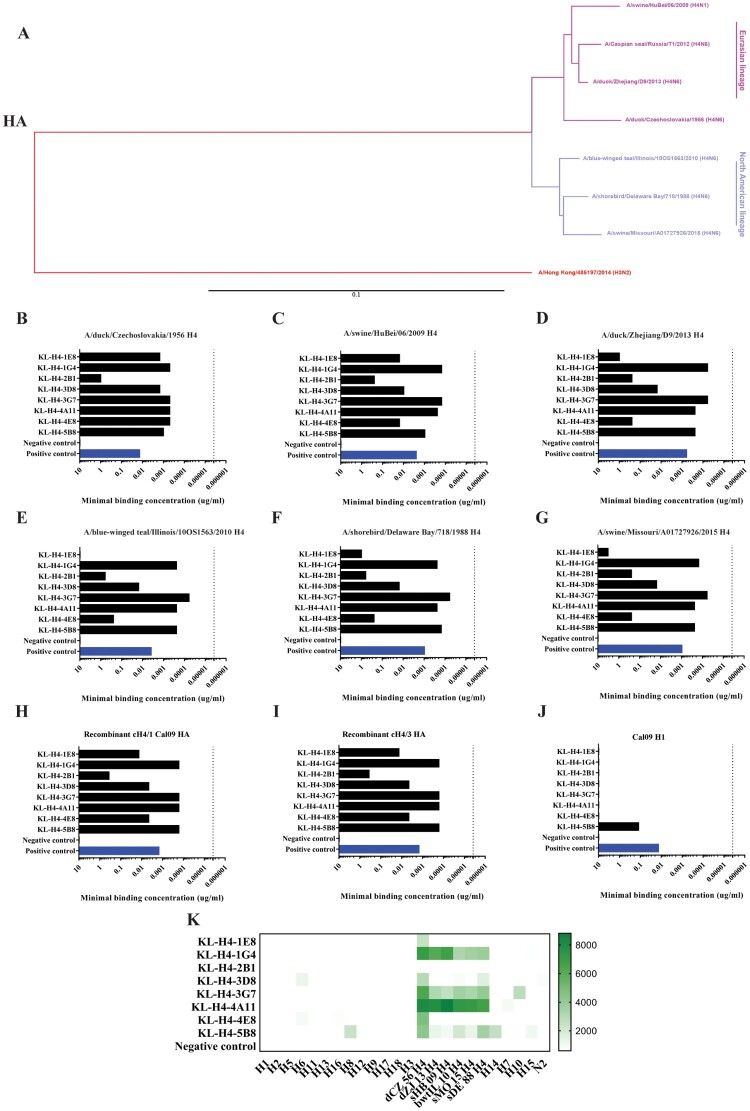
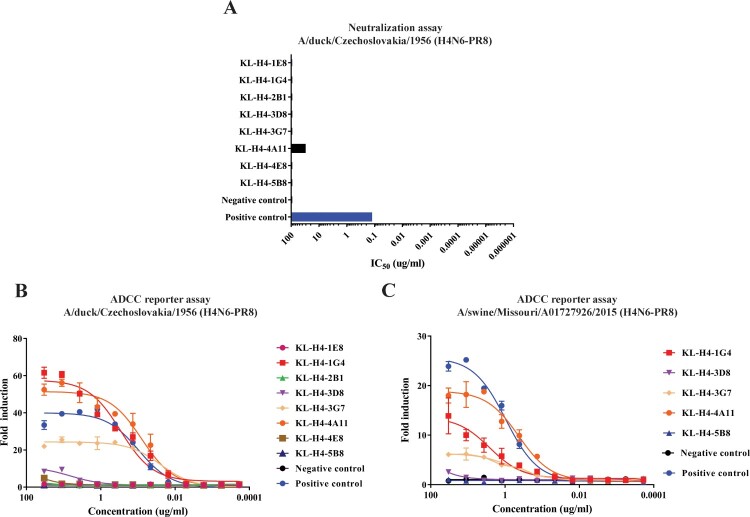
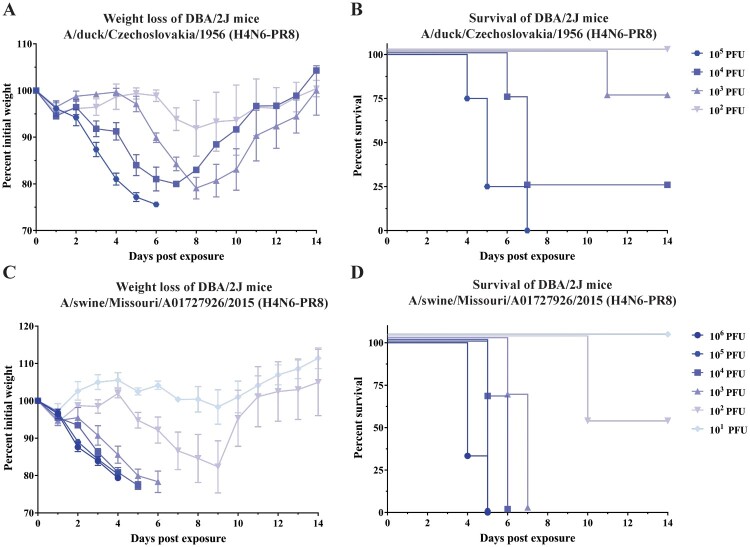
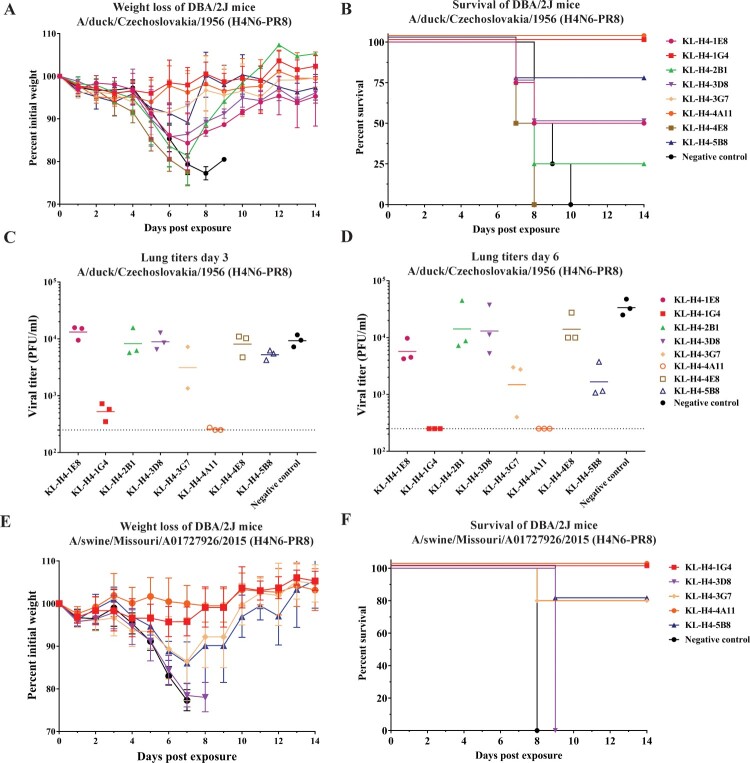
Influenza viruses of the H4 subtype are widespread in wild birds, circulate in domestic poultry, readily infect mammals, and tolerate the insertion of a polybasic cleavage site. In addition, serological evidence suggests that humans working with poultry are exposed to these viruses. While H4 viruses are not of immediate pandemic concern, there is a lack of knowledge regarding their antigenicity. In order to study viruses of the H4 subtype, we generated and characterized a panel of antibodies that bind a wide variety of H4 hemagglutinins from avian and swine isolates of both the Eurasian and North American lineage. We further characterized these antibodies using novel recombinant H4N6 viruses that were found to be lethal in DBA/2J mice. Non-neutralizing antibodies, which had activity in an antibody dependent cell-mediated cytotoxicity reporter assay in vitro, protected mice against challenge in vivo, highlighting the importance of effector functions. Our data suggest a high degree of antigenic conservation of the H4 hemagglutinin.
Keywords: H4N6; Influenza virus; avian influenza; hemagglutinin; monoclonal antibodies.
Conflict of interest statement
The Icahn School of Medicine at Mount Sinai has filed patent applications regarding influenza virus vaccines and Florian Krammer is named as inventor on several of these applications.
Figures
Similar articles
-
In Vitro Neutralization Is Not Predictive of Prophylactic Efficacy of Broadly Neutralizing Monoclonal Antibodies CR6261 and CR9114 against Lethal H2 Influenza Virus Challenge in Mice.J Virol. 2017 Nov 30;91(24):e01603-17. doi: 10.1128/JVI.01603-17. Print 2017 Dec 15. J Virol. 2017. PMID: 29046448 Free PMC article.
-
Hemagglutinin Stalk- and Neuraminidase-Specific Monoclonal Antibodies Protect against Lethal H10N8 Influenza Virus Infection in Mice.J Virol. 2015 Oct 28;90(2):851-61. doi: 10.1128/JVI.02275-15. Print 2016 Jan 15. J Virol. 2015. PMID: 26512088 Free PMC article.
-
Cross-reactive mouse monoclonal antibodies raised against the hemagglutinin of A/Shanghai/1/2013 (H7N9) protect against novel H7 virus isolates in the mouse model.Emerg Microbes Infect. 2018 Jun 20;7(1):110. doi: 10.1038/s41426-018-0115-0. Emerg Microbes Infect. 2018. PMID: 29925896 Free PMC article.
-
One step closer to universal influenza epitopes.Expert Rev Anti Infect Ther. 2009 Aug;7(6):687-90. doi: 10.1586/eri.09.48. Expert Rev Anti Infect Ther. 2009. PMID: 19681695 Review.
-
Influenza virus glycoprotein-reactive human monoclonal antibodies.Microbes Infect. 2020 Jul-Aug;22(6-7):263-271. doi: 10.1016/j.micinf.2020.06.003. Epub 2020 Jun 19. Microbes Infect. 2020. PMID: 32569735 Free PMC article. Review.
Cited by
-
A serological assay to detect SARS-CoV-2 seroconversion in humans.medRxiv [Preprint]. 2020 Apr 16:2020.03.17.20037713. doi: 10.1101/2020.03.17.20037713. medRxiv. 2020. Update in: Nat Med. 2020 Jul;26(7):1033-1036. doi: 10.1038/s41591-020-0913-5. PMID: 32511441 Free PMC article. Updated. Preprint.
-
Murine Monoclonal Antibodies against the Receptor Binding Domain of SARS-CoV-2 Neutralize Authentic Wild-Type SARS-CoV-2 as Well as B.1.1.7 and B.1.351 Viruses and Protect In Vivo in a Mouse Model in a Neutralization-Dependent Manner.mBio. 2021 Aug 31;12(4):e0100221. doi: 10.1128/mBio.01002-21. Epub 2021 Jul 27. mBio. 2021. PMID: 34311574 Free PMC article.
-
Assessment of a quadrivalent nucleoside-modified mRNA vaccine that protects against group 2 influenza viruses.Proc Natl Acad Sci U S A. 2022 Nov 8;119(45):e2206333119. doi: 10.1073/pnas.2206333119. Epub 2022 Nov 2. Proc Natl Acad Sci U S A. 2022. PMID: 36322769 Free PMC article.
-
Enhanced Potency of a Broad H7N9-Neutralizing Antibody HNIgGA6 Through Structure-Based Design.Front Microbiol. 2020 Jun 19;11:1313. doi: 10.3389/fmicb.2020.01313. eCollection 2020. Front Microbiol. 2020. PMID: 32636820 Free PMC article.
-
Vaccination With Viral Vectors Expressing Chimeric Hemagglutinin, NP and M1 Antigens Protects Ferrets Against Influenza Virus Challenge.Front Immunol. 2019 Aug 21;10:2005. doi: 10.3389/fimmu.2019.02005. eCollection 2019. Front Immunol. 2019. PMID: 31497029 Free PMC article.
References
-
- Krammer F. Emerging influenza viruses and the prospect of a universal influenza virus vaccine. Biotechnol J. 2015;10:690–701. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources