

Endoplasmic reticulum-localized UBC34 interaction with lignin repressors MYB221 and MYB156 regulates the transactivity of the transcription factors in Populus tomentosa
- PMID: 30866808
- PMCID: PMC6416899
- DOI: 10.1186/s12870-019-1697-y
Endoplasmic reticulum-localized UBC34 interaction with lignin repressors MYB221 and MYB156 regulates the transactivity of the transcription factors in Populus tomentosa
Abstract
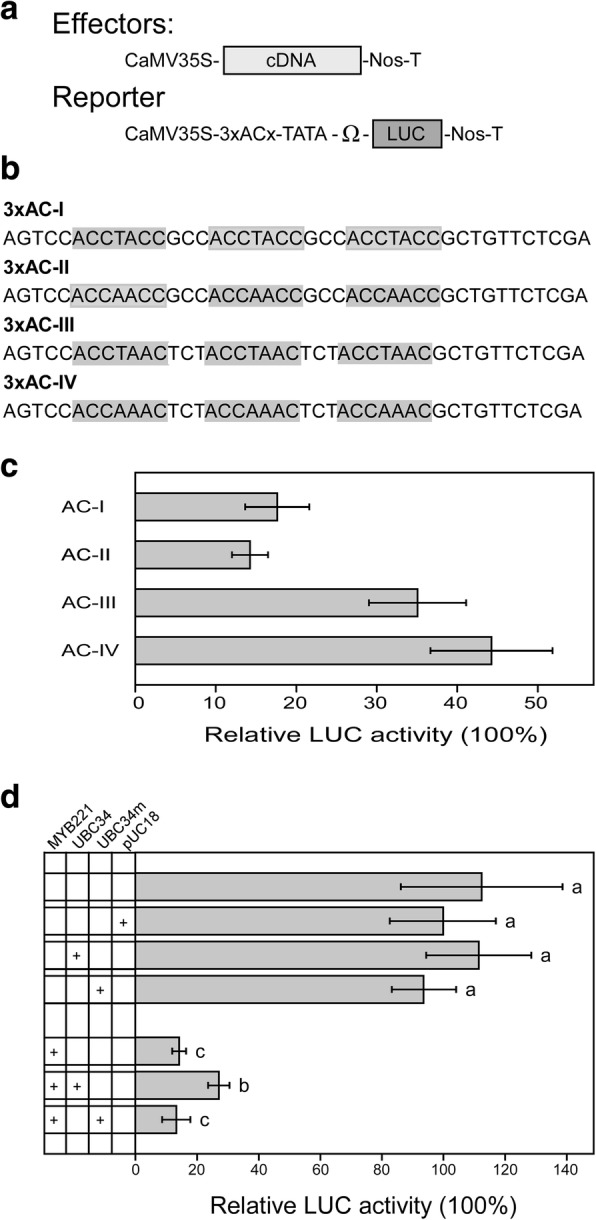
Background: Regulation of lignin biosynthesis is known to occur at the level of transcription factors (TFs), of which R2R3-MYB family members have been proposed to play a central role via the AC cis-elements. Despite the important roles of TFs in lignin biosynthesis, the post-translational regulation of these TFs, particularly their ubiquitination regulation, has not been thoroughly explored.
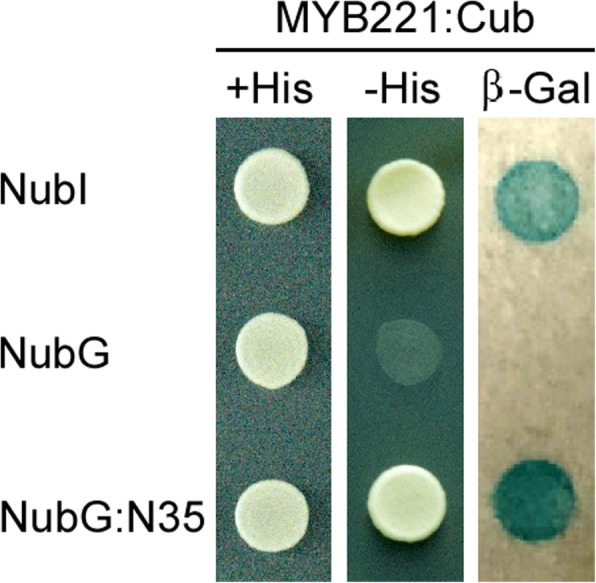
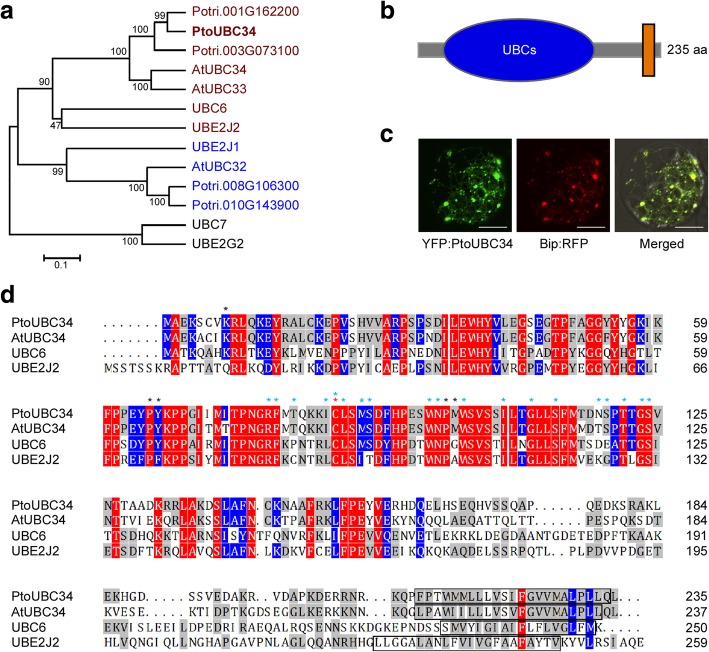
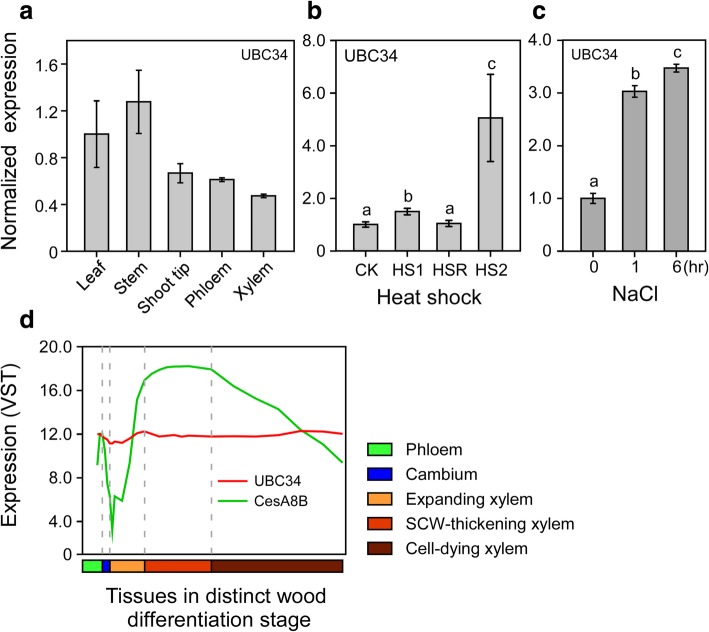
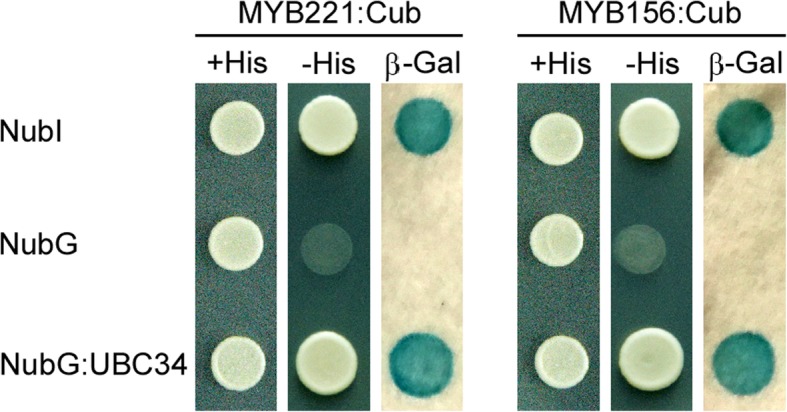
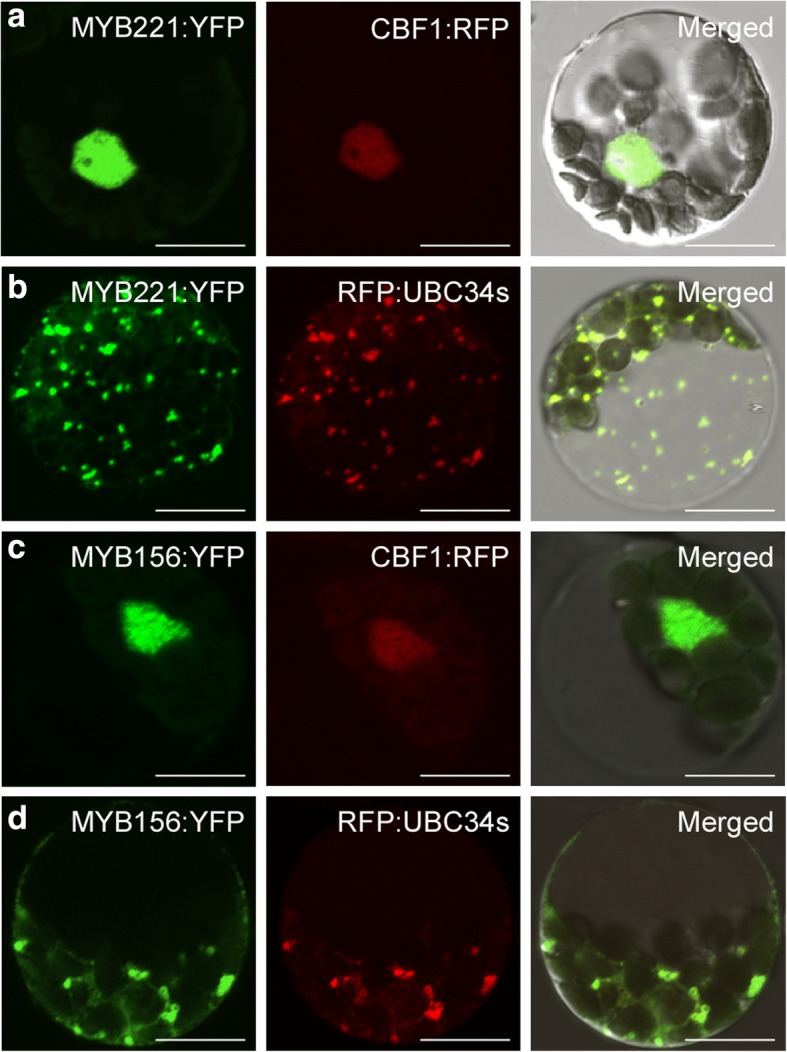
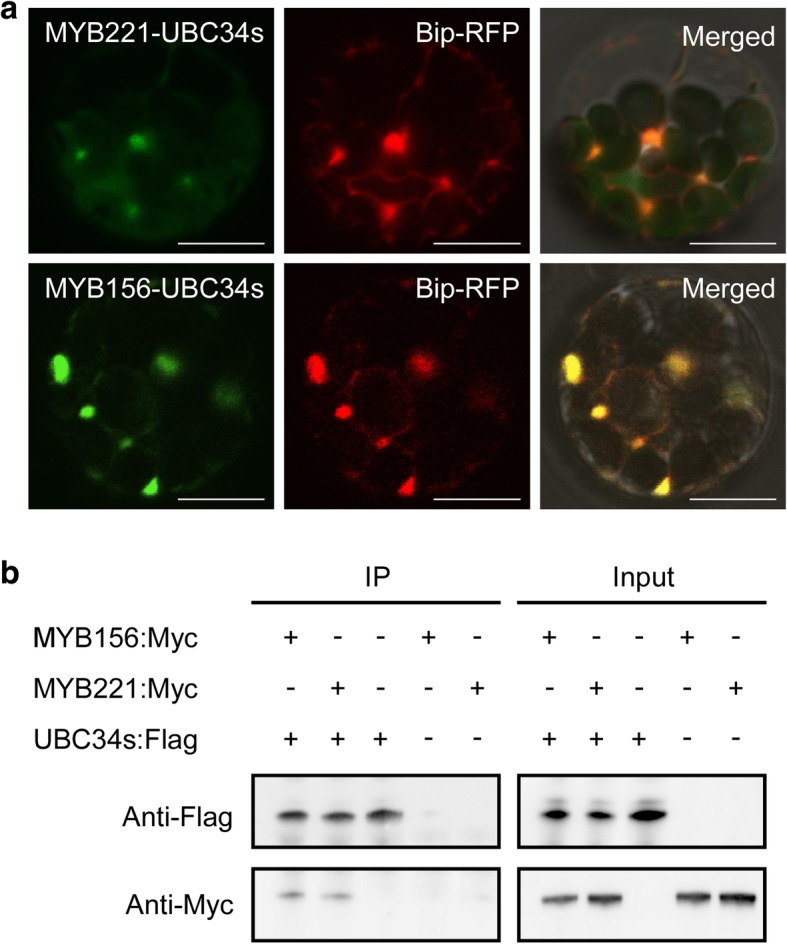
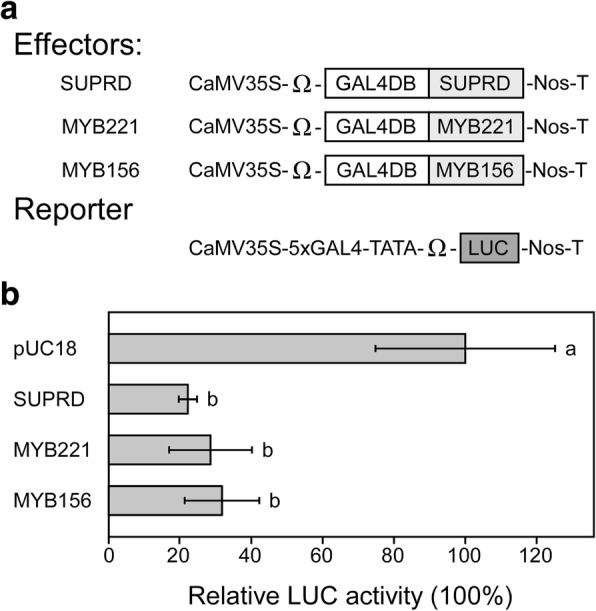
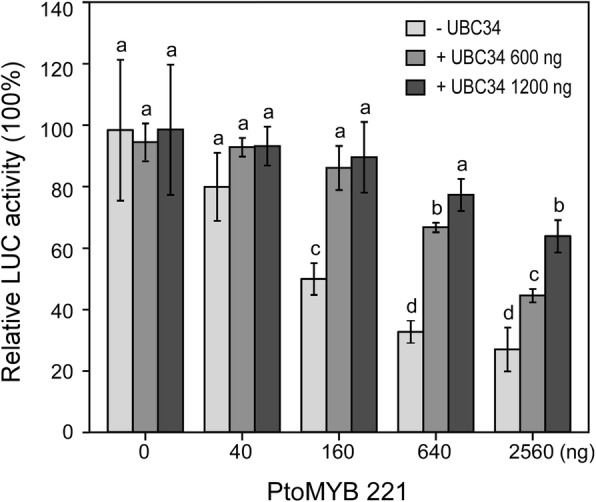
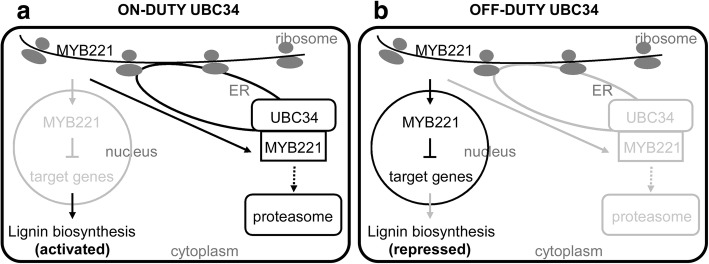
Results: We describe the discovery of a Populus tomentosa E2 ubiquitin-conjugating enzyme 34 (PtoUBC34), which is involved in the post-translational regulation of transactivation activity of lignin-associated transcriptional repressors PtoMYB221 and PtoMYB156. PtoUBC34 is localized at the endoplasmic reticulum (ER) membrane where it interacts with transcriptional repressors PtoMYB221 and PtoMYB156. This specific interaction allows for the translocation of TFs PtoMYB221 and PtoMYB156 to the ER and reduces their repression activity in a PtoUBC34 abundance-dependent manner. By taking a molecular biology approach with quantitative real-time polymerase chain reaction (qRT-PCR) analysis, we found that PtoUBC34 is expressed in all aboveground tissues of trees in P. tomentosa, and in particular, it is ubiquitous in all distinct differentiation stages across wood formation, including phloem differentiation, cambium maintaining, early and developing xylem differentiation, secondary cell wall thickening, and programmed cell death. Additionally, we discovered that PtoUBC34 is induced by treatment with sodium chloride and heat shock.
Conclusions: Our data suggest a possible mechanism by which lignin biosynthesis is regulated by ER-localized PtoUBC34 in poplar, probably through the ER-associated degradation (ERAD) of lignin-associated repressors PtoMYB221 and PtoMYB156.
Keywords: ER-associated degradation; Lignin biosynthesis; PtoMYB156; PtoMYB221; PtoUBC34; Transcriptional repressor.
Conflict of interest statement
Ethics approval and consent to participate
The plant materials were collected from the Research base of Beijing Academy of Agriculture and Forestry Sciences, Beijing, China. The experimental research on plants, including collection of plant material, was complied with the institutional, national, or international guidelines. No specific permits were required by the scientific research institute to use these materials.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Weng JK, Chapple C. The origin and evolution of lignin biosynthesis. New Phytol. 2010;187(2):273–285. - PubMed
-
- Boerjan W, Ralph J, Baucher M. Lignin biosynthesis. Annu Rev Plant Biol. 2003;54:519–546. - PubMed
-
- Pilate G, Guiney E, Holt K, Petit-Conil M, Lapierre C, Leple JC, Pollet B, Mila I, Webster EA, Marstorp HG, et al. Field and pulping performances of transgenic trees with altered lignification. Nat Biotechnol. 2002;20(6):607–612. - PubMed
-
- Himmel ME, Ding SY, Johnson DK, Adney WS, Nimlos MR, Brady JW, Foust TD. Biomass recalcitrance: engineering plants and enzymes for biofuels production. Science. 2007;315(5813):804–807. - PubMed
-
- Baucher M, Halpin C, Petit-Conil M, Boerjan W. Lignin: genetic engineering and impact on pulping. Crit Rev Biochem Mol Biol. 2003;38(4):305–350. - PubMed
MeSH terms
Substances
Grants and funding
- 31770708/National Natural Science Foundation of China
- 31270593/National Natural Science Foundation of China
- 2016YFD0600104/National Key R&D Program of China
- KJCX20170203/Science Foundation of the Beijing Academy of Agriculture and Forestry Sciences
- 2018ZX0802002/National Key Program on Transgenic Research of China
LinkOut - more resources
Full Text Sources
