

A scalable solution for isolating human multipotent clinical-grade neural stem cells from ES precursors
- PMID: 30867054
- PMCID: PMC6417180
- DOI: 10.1186/s13287-019-1163-7
A scalable solution for isolating human multipotent clinical-grade neural stem cells from ES precursors
Abstract
Background: A well-characterized method has not yet been established to reproducibly, efficiently, and safely isolate large numbers of clinical-grade multipotent human neural stem cells (hNSCs) from embryonic stem cells (hESCs). Consequently, the transplantation of neurogenic/gliogenic precursors into the CNS for the purpose of cell replacement or neuroprotection in humans with injury or disease has not achieved widespread testing and implementation.
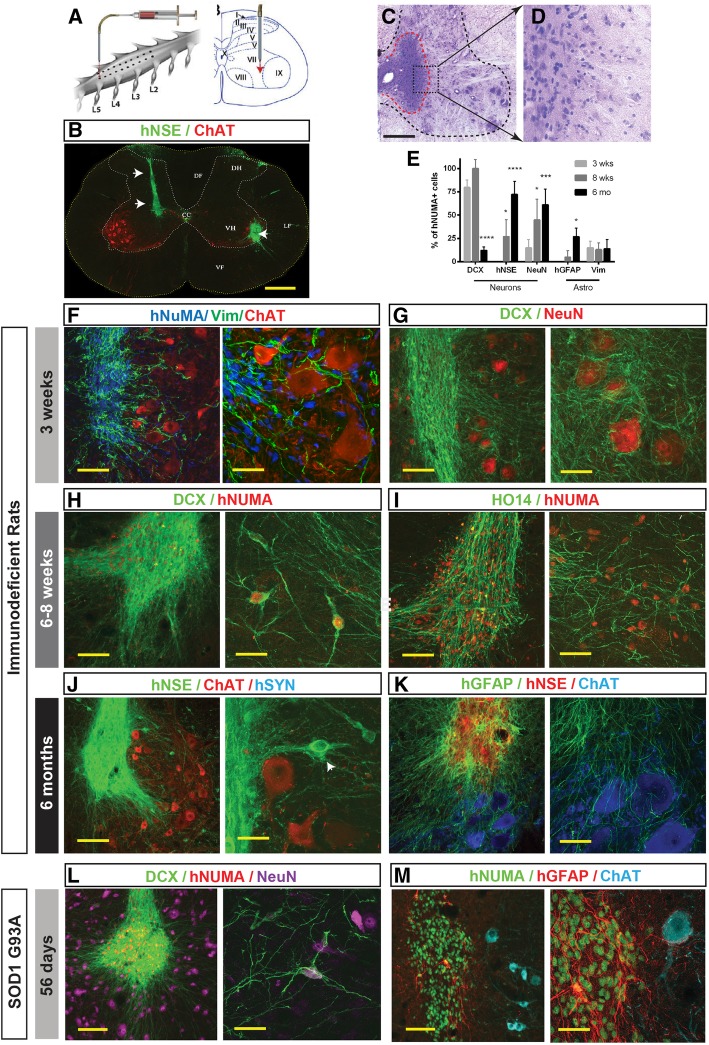
Methods: Here, we establish an approach for the in vitro isolation of a highly expandable population of hNSCs using the manual selection of neural precursors based on their colony morphology (CoMo-NSC). The purity and NSC properties of established and extensively expanded CoMo-NSC were validated by expression of NSC markers (flow cytometry, mRNA sequencing), lack of pluripotent markers and by their tumorigenic/differentiation profile after in vivo spinal grafting in three different animal models, including (i) immunodeficient rats, (ii) immunosuppressed ALS rats (SOD1G93A), or (iii) spinally injured immunosuppressed minipigs.
Results: In vitro analysis of established CoMo-NSCs showed a consistent expression of NSC markers (Sox1, Sox2, Nestin, CD24) with lack of pluripotent markers (Nanog) and stable karyotype for more than 15 passages. Gene profiling and histology revealed that spinally grafted CoMo-NSCs differentiate into neurons, astrocytes, and oligodendrocytes over a 2-6-month period in vivo without forming neoplastic derivatives or abnormal structures. Moreover, transplanted CoMo-NSCs formed neurons with synaptic contacts and glia in a variety of host environments including immunodeficient rats, immunosuppressed ALS rats (SOD1G93A), or spinally injured minipigs, indicating these cells have favorable safety and differentiation characteristics.
Conclusions: These data demonstrate that manually selected CoMo-NSCs represent a safe and expandable NSC population which can effectively be used in prospective human clinical cell replacement trials for the treatment of a variety of neurodegenerative disorders, including ALS, stroke, spinal traumatic, or spinal ischemic injury.
Keywords: Amyotrophic lateral sclerosis (ALS); Bioinformatic tools to study xenografts; Human embryonic stem cell (hESC); Neural stem cell (NSC); Spinal cord; Spinal traumatic injury.
Conflict of interest statement
Ethics approval and consent to participate
Animal studies
All animal studies were approved by the University of California, San Diego Institutional Animal Care and Use Committee (Protocol No.: S01193) and by the Institutional Animal Care and Use Committee of the Czech Academy of Sciences and were in compliance with The Association for Assessment of Laboratory Animal Care guidelines for animal use.
Use of human ES lines
This study was approved by the University of California, San Diego (UCSD) Internal Review Board (IRB), (approval ID#101323ZX).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures





Similar articles
-
Survival and differentiation of human embryonic stem cell-derived neural precursors grafted spinally in spinal ischemia-injured rats or in naive immunosuppressed minipigs: a qualitative and quantitative study.Cell Transplant. 2012;21(12):2603-19. doi: 10.3727/096368912X653200. Epub 2012 Aug 10. Cell Transplant. 2012. PMID: 22889456
-
Grafted Subventricular Zone Neural Stem Cells Display Robust Engraftment and Similar Differentiation Properties and Form New Neurogenic Niches in the Young and Aged Hippocampus.Stem Cells Transl Med. 2016 Sep;5(9):1204-15. doi: 10.5966/sctm.2015-0270. Epub 2016 May 18. Stem Cells Transl Med. 2016. PMID: 27194744 Free PMC article.
-
Cell-surface marker signatures for the isolation of neural stem cells, glia and neurons derived from human pluripotent stem cells.PLoS One. 2011 Mar 2;6(3):e17540. doi: 10.1371/journal.pone.0017540. PLoS One. 2011. PMID: 21407814 Free PMC article.
-
Stem cell repair of central nervous system injury.J Neurosci Res. 2002 Jun 1;68(5):501-10. doi: 10.1002/jnr.10240. J Neurosci Res. 2002. PMID: 12111840 Review.
-
Human neural stem cells genetically modified for brain repair in neurological disorders.Neuropathology. 2004 Sep;24(3):159-71. doi: 10.1111/j.1440-1789.2004.00552.x. Neuropathology. 2004. PMID: 15484694 Review.
Cited by
-
Routes and methods of neural stem cells injection in cerebral ischemia.Ibrain. 2023 Aug 6;9(3):326-339. doi: 10.1002/ibra.12122. eCollection 2023 Fall. Ibrain. 2023. PMID: 37786754 Free PMC article. Review.
-
Xeno-free induced pluripotent stem cell-derived neural progenitor cells for in vivo applications.J Transl Med. 2022 Sep 16;20(1):421. doi: 10.1186/s12967-022-03610-5. J Transl Med. 2022. PMID: 36114512 Free PMC article.
-
Derivation of Sendai-Virus-Reprogrammed Human iPSCs-Neuronal Precursors: In Vitro and In Vivo Post-grafting Safety Characterization.Cell Transplant. 2023 Jan-Dec;32:9636897231163232. doi: 10.1177/09636897231163232. Cell Transplant. 2023. PMID: 36959733 Free PMC article.
-
How neural stem cell therapy promotes brain repair after stroke.Stem Cell Reports. 2025 Jun 10;20(6):102507. doi: 10.1016/j.stemcr.2025.102507. Epub 2025 May 22. Stem Cell Reports. 2025. PMID: 40409262 Free PMC article. Review.
-
Effects of all-trans and 9-cis retinoic acid on differentiating human neural stem cells in vitro.Toxicology. 2023 Mar 15;487:153461. doi: 10.1016/j.tox.2023.153461. Epub 2023 Feb 16. Toxicology. 2023. PMID: 36805303 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
