

Postnatal development and maturation of layer 1 in the lateral prefrontal cortex and its disruption in autism
- PMID: 30867066
- PMCID: PMC6417186
- DOI: 10.1186/s40478-019-0684-8
Postnatal development and maturation of layer 1 in the lateral prefrontal cortex and its disruption in autism
Abstract
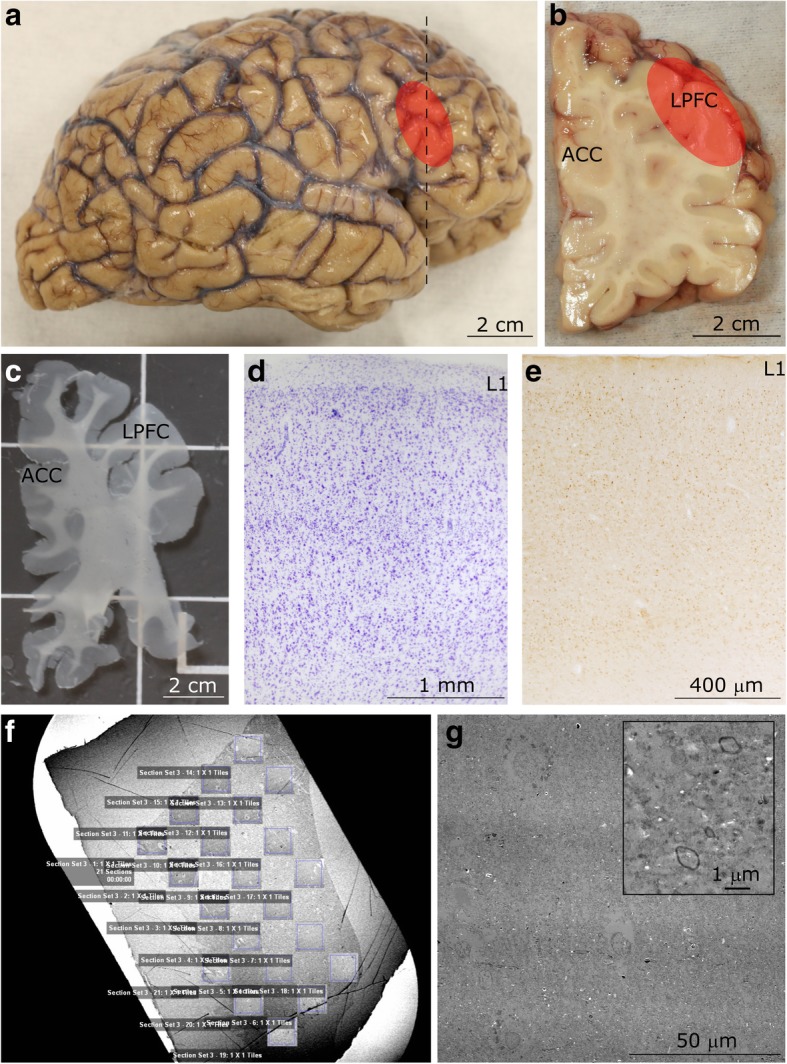
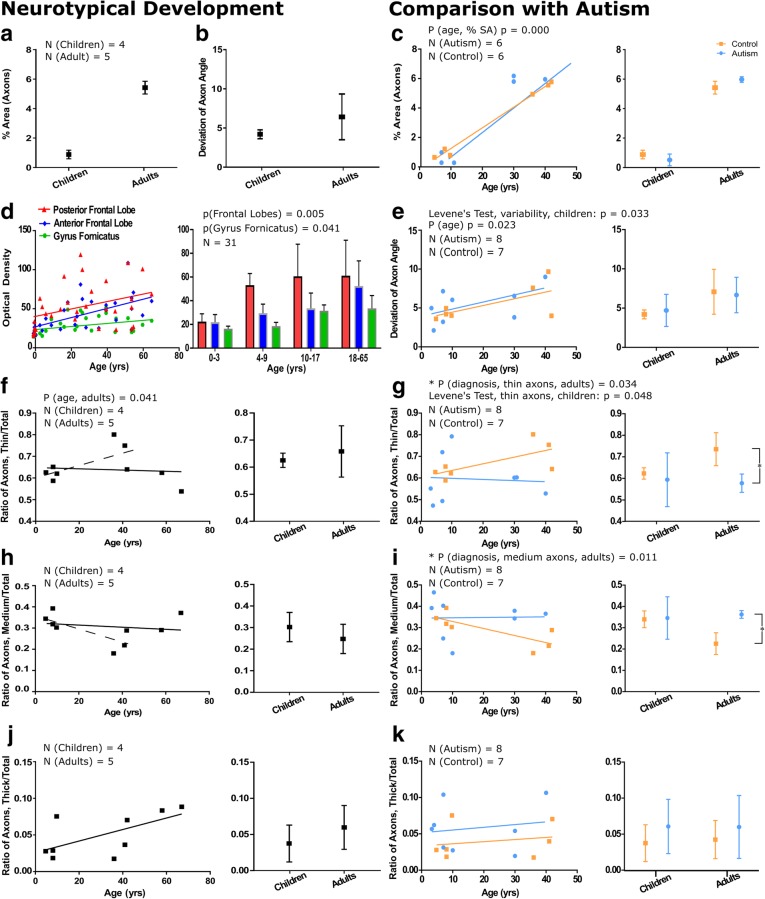
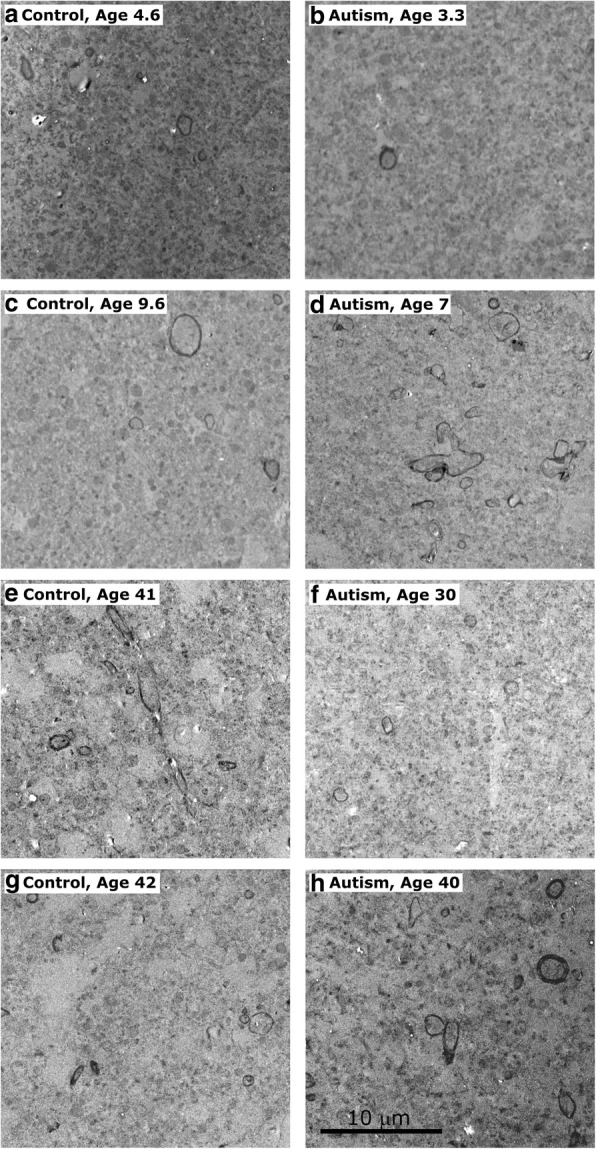
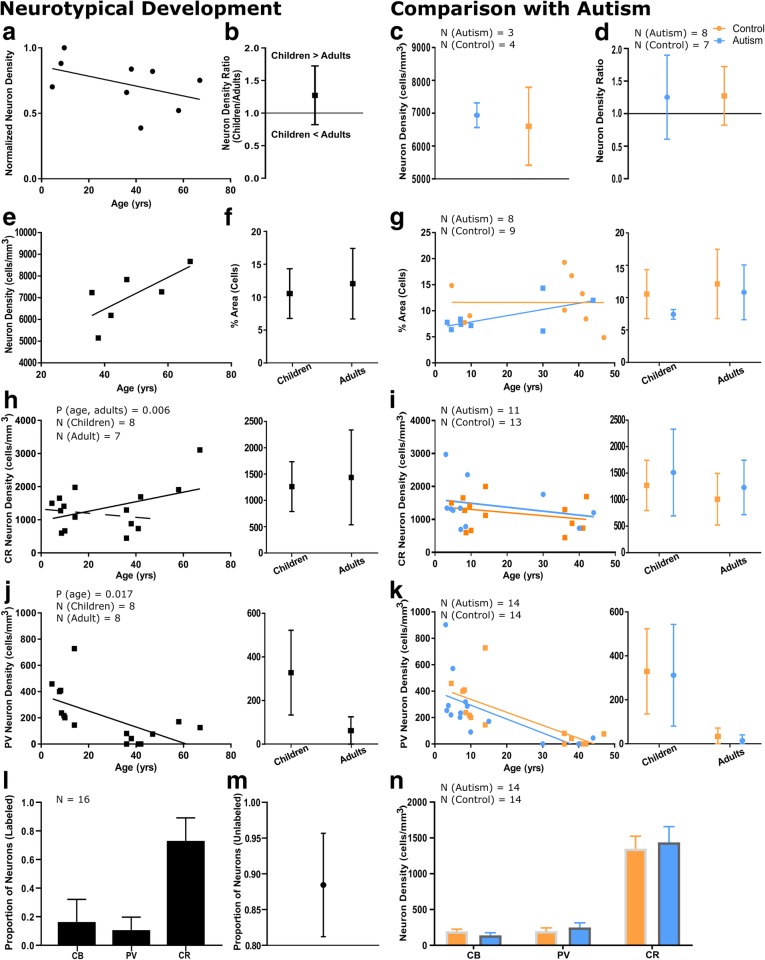
Autism is a neurodevelopmental connectivity disorder characterized by cortical network disorganization and imbalance in excitation/inhibition. However, little is known about the development of autism pathology and the disruption of laminar-specific excitatory and inhibitory cortical circuits. To begin to address these issues, we examined layer 1 of the lateral prefrontal cortex (LPFC), an area with prolonged development and maturation that is affected in autism. We focused on layer 1 because it contains a distinctive, diverse population of interneurons and glia, receives input from feedback and neuromodulatory pathways, and plays a critical role in the development, maturation, and function of the cortex. We used unbiased quantitative methods at high resolution to study the morphology, neurochemistry, distribution, and density of neurons and myelinated axons in post-mortem brain tissue from children and adults with and without autism. We cross-validated our findings through comparisons with neighboring anterior cingulate cortices and optimally-fixed non-human primate tissue. In neurotypical controls we found an increase in the density of myelinated axons from childhood to adulthood. Neuron density overall declined with age, paralleled by decreased density of inhibitory interneurons labeled by calretinin (CR), calbindin (CB), and parvalbumin (PV). Importantly, we found PV neurons in layer 1 of typically developing children, previously detected only perinatally. In autism there was disorganization of cortical networks within layer 1: children with autism had increased variability in the trajectories and thickness of myelinated axons in layer 1, while adults with autism had a reduction in the relative proportion of thin axons. Neurotypical postnatal changes in layer 1 of LPFC likely underlie refinement of cortical activity during maturation of cortical networks involved in cognition. Our findings suggest that disruption of the maturation of feedback pathways, rather than interneurons in layer 1, has a key role in the development of imbalance between excitation and inhibition in autism.
Keywords: Anterior cingulate cortex; Autism neuropathology; Feedback pathways; Inhibitory neuron; Laminar architecture; Postnatal axon myelination.
Conflict of interest statement
Ethics approval and consent to participate
The use of human post-mortem tissue for this study was approved by the Institutional Review Board (IRB) of Boston University. Experiments and procedures with animals were designed to minimize animal suffering and reduce the number of animals used. Detailed protocols of the procedures were approved by the Institutional Animal Care and Use Committee (IACUC) at Harvard Medical School and Boston University School of Medicine in accordance with NIH guidelines (DHEW Publication no. [NIH] 80–22, revised 1996, Office of Science and Health Reports, DRR/NIH, Bethesda, Maryland, United States).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
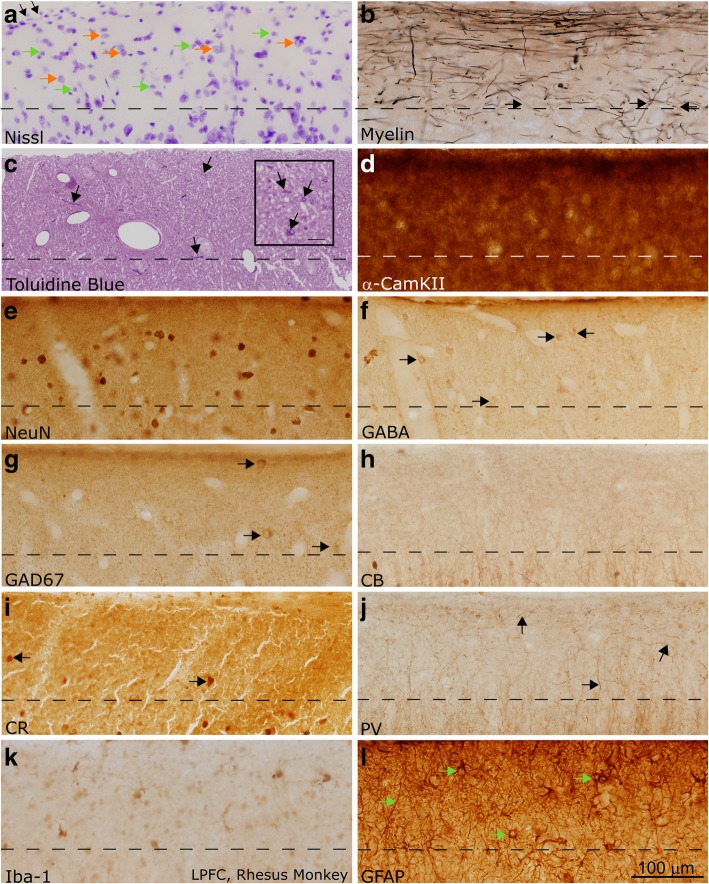
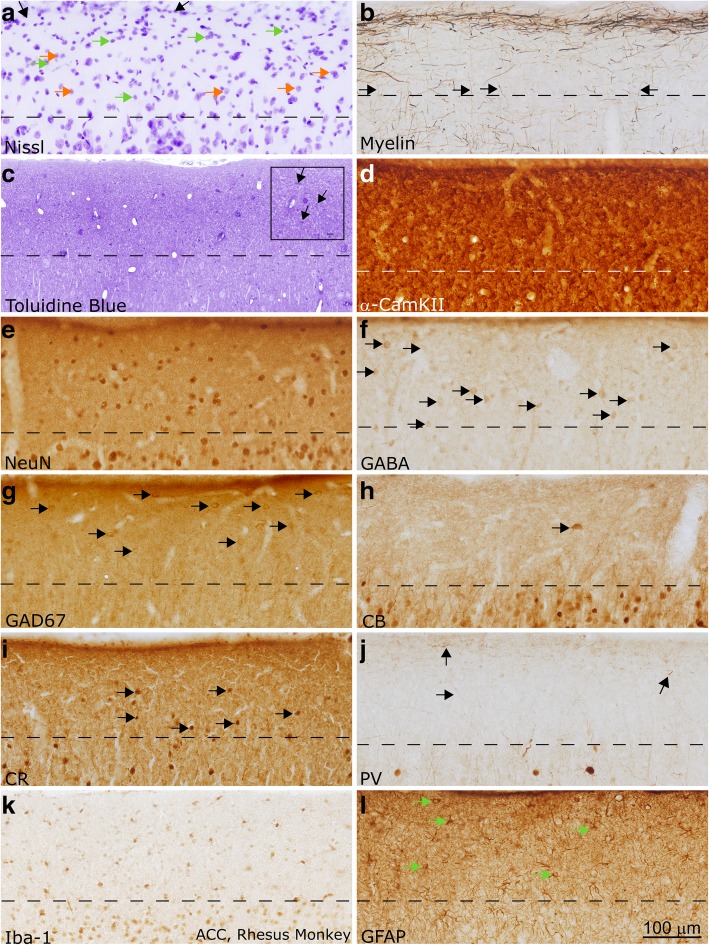
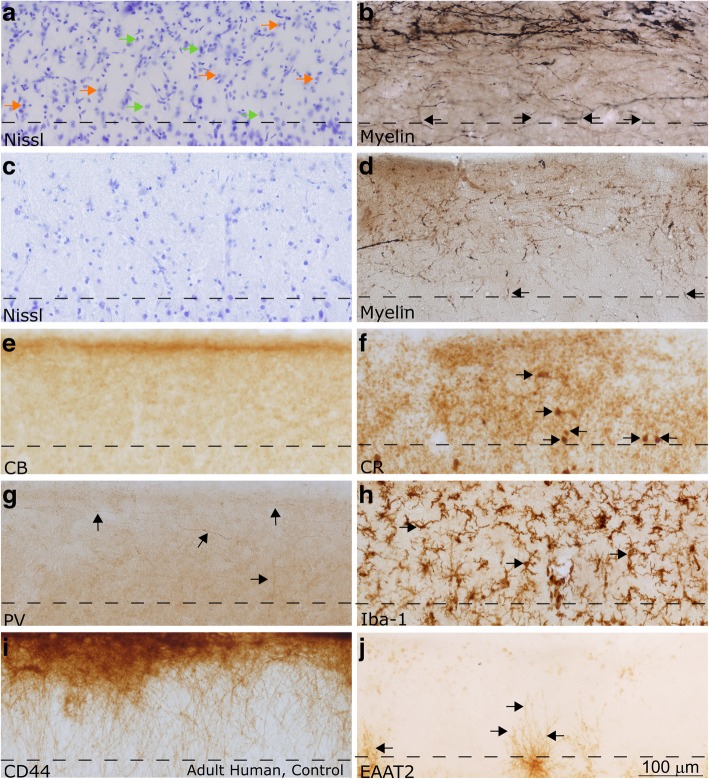
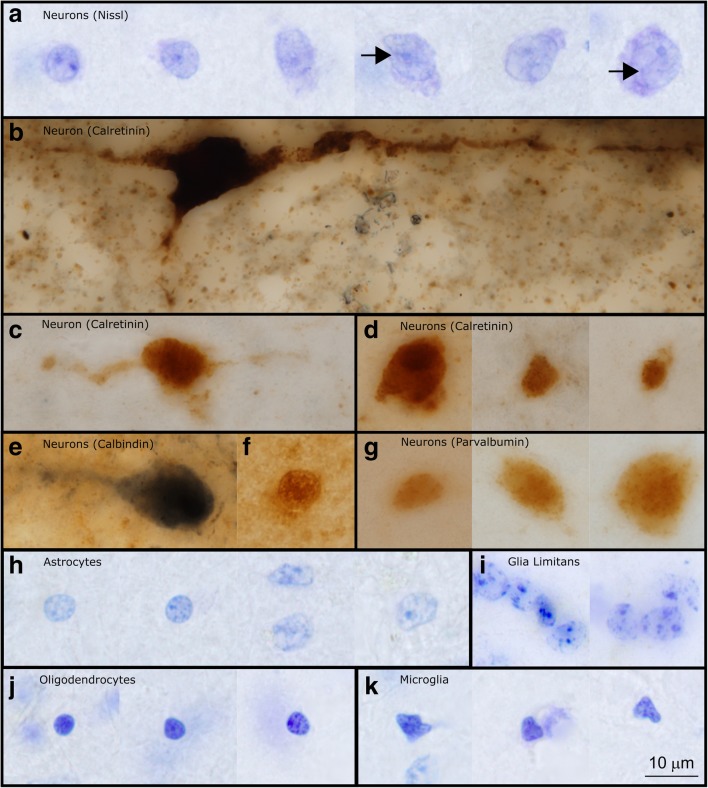
References
-
- Barbaria EM, Kohl B, Buhren BA, Hasenpusch-Theil K, Kruse F, Kury P, Martini R, Muller HW. The alpha-chemokine CXCL14 is up-regulated in the sciatic nerve of a mouse model of Charcot-Marie-tooth disease type 1A and alters myelin gene expression in cultured Schwann cells. Neurobiol Dis. 2009;33:448–458. doi: 10.1016/j.nbd.2008.11.014. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
