

Diverse fate of ubiquitin chain moieties: The proximal is degraded with the target, and the distal protects the proximal from removal and recycles
- PMID: 30867293
- PMCID: PMC6475391
- DOI: 10.1073/pnas.1822148116
Diverse fate of ubiquitin chain moieties: The proximal is degraded with the target, and the distal protects the proximal from removal and recycles
Abstract
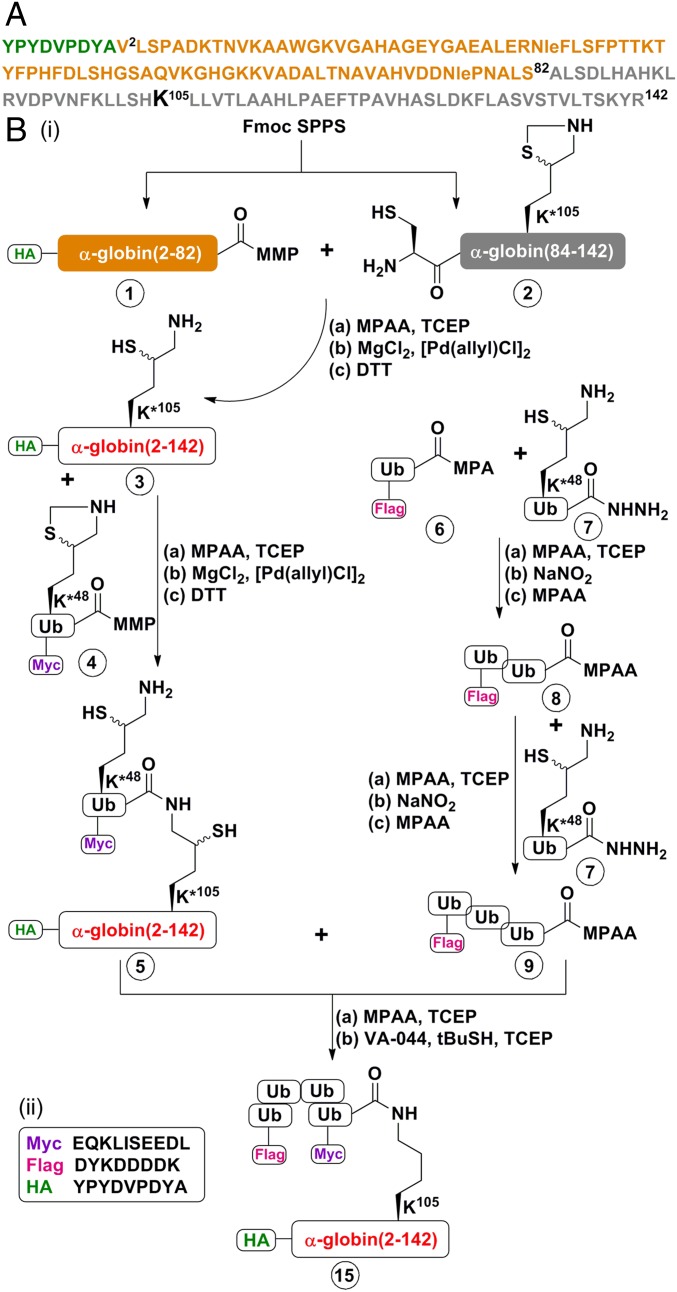
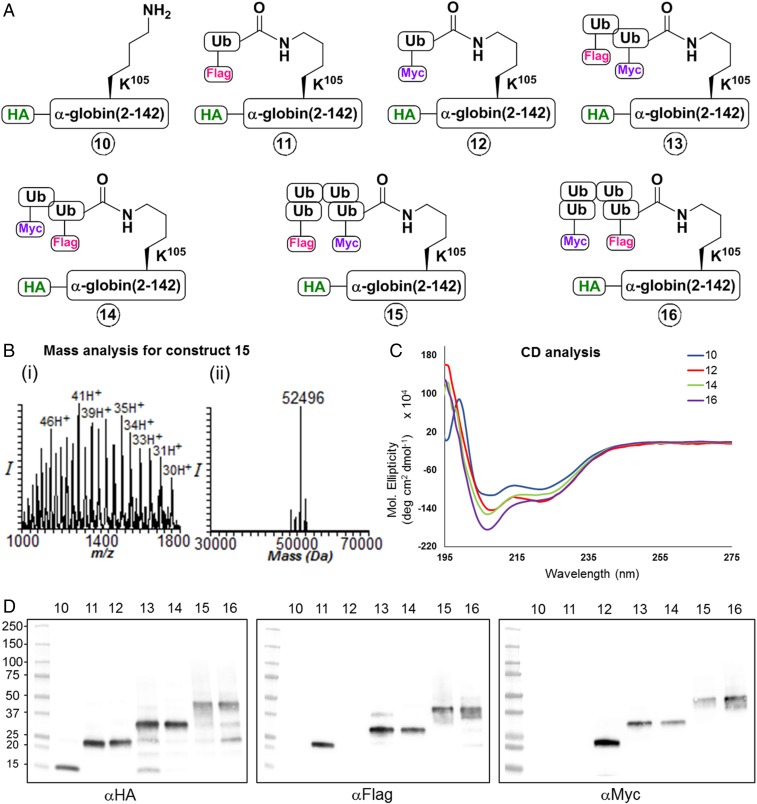
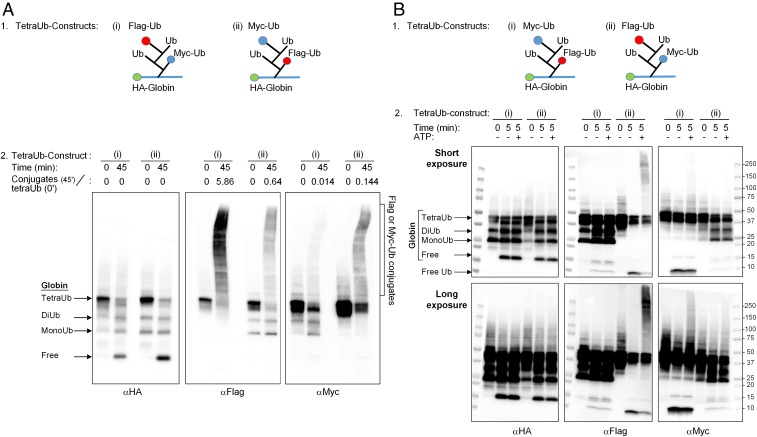
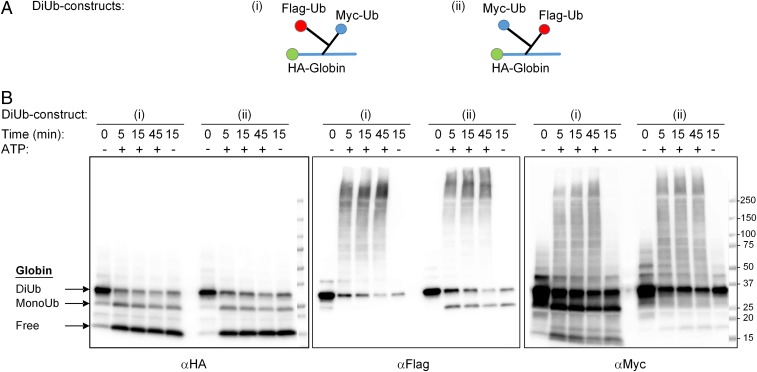
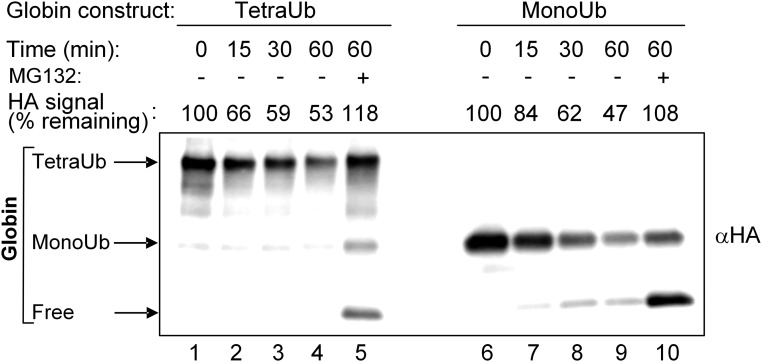
One of the enigmas in the ubiquitin (Ub) field is the requirement for a poly-Ub chain as a proteasomal targeting signal. The canonical chain appears to be longer than the distance between the two Ub-binding proteasomal receptors. Furthermore, genetic manipulation has shown that one receptor subunit is sufficient, which suggests that a single Ub can serve as a degradation signal. To shed light on this mystery, we chemically synthesized tetra-Ub, di-Ub (K48-based), and mono-Ub adducts of HA-α-globin, where the distal or proximal Ub moieties were tagged differentially with either Myc or Flag. When incubated in a crude cell extract, the distal Ub moiety in the tetra-Ub adduct was mostly removed by deubiquitinating enzymes (DUBs) and reconjugated to other substrates in the extract. In contrast, the proximal moiety was most likely degraded with the substrate. The efficacy of degradation was proportionate to the chain length; while tetra-Ub globin was an efficient substrate, with mono-Ub globin, we observed rapid removal of the Ub moiety with almost no degradation of the free globin. Taken together, these findings suggest that the proximal moieties are necessary for securing the association of the substrate with the proteasome along the proteolytic process, whereas the distal moieties are important in protecting the proximal moieties from premature deubiquitination. Interestingly, when the same experiment was carried out using purified 26S proteasome, mono- and tetra-Ub globin were similarly degraded, highlighting the roles of the entire repertoire of cellular DUBs in regulating the degradation of proteasomal substrates.
Keywords: 26S proteasome; chemical synthesis; deubiquitination; protein degradation; ubiquitination.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Comment in
-
Synthetic ubiquitinated proteins meet the proteasome: Distinct roles of ubiquitin in a chain.Proc Natl Acad Sci U S A. 2019 Apr 16;116(16):7614-7616. doi: 10.1073/pnas.1903405116. Epub 2019 Mar 29. Proc Natl Acad Sci U S A. 2019. PMID: 30926663 Free PMC article. No abstract available.
References
-
- Mali SM, Singh SK, Eid E, Brik A. Ubiquitin signaling: Chemistry comes to the rescue. J Am Chem Soc. 2017;139:4971–4986. - PubMed
-
- Komander D, Rape M. The ubiquitin code. Annu Rev Biochem. 2012;81:203–229. - PubMed
-
- Kulathu Y, Komander D. Atypical ubiquitylation—The unexplored world of polyubiquitin beyond Lys48 and Lys63 linkages. Nat Rev Mol Cell Biol. 2012;13:508–523. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
