

A Novel Fluorescent and Bioluminescent Bireporter Influenza A Virus To Evaluate Viral Infections
- PMID: 30867298
- PMCID: PMC6498038
- DOI: 10.1128/JVI.00032-19
A Novel Fluorescent and Bioluminescent Bireporter Influenza A Virus To Evaluate Viral Infections
Abstract
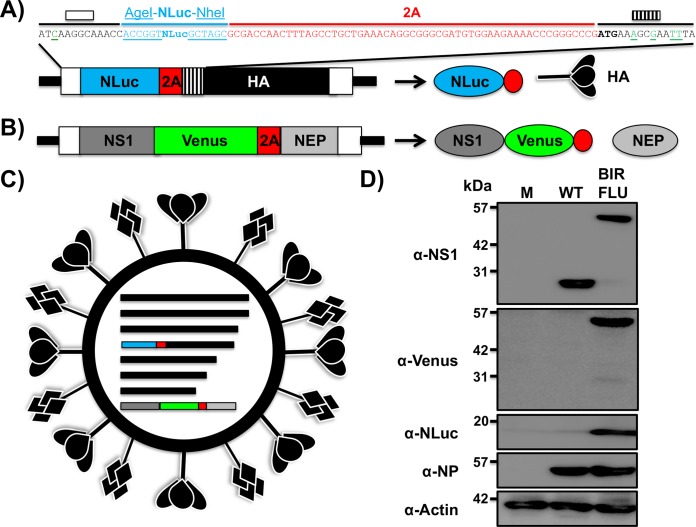
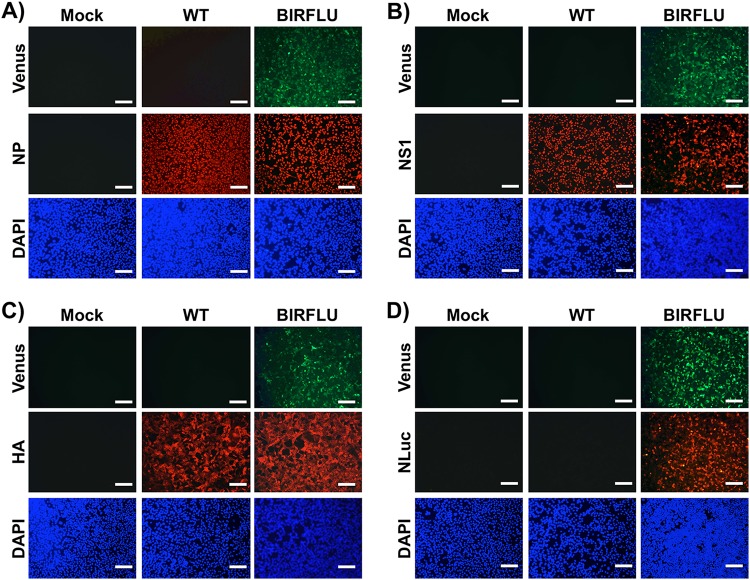
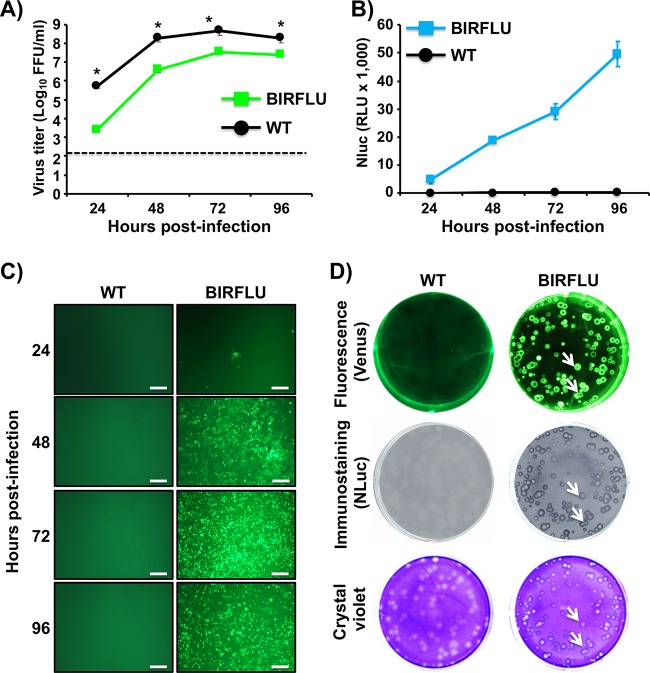
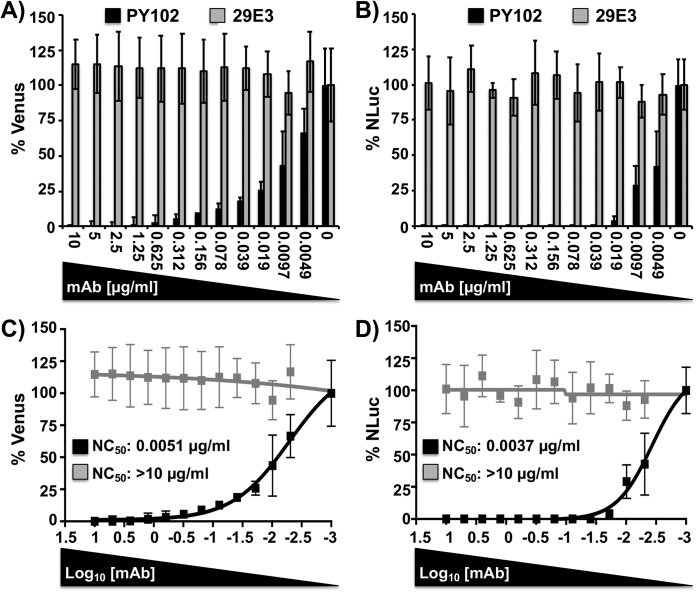
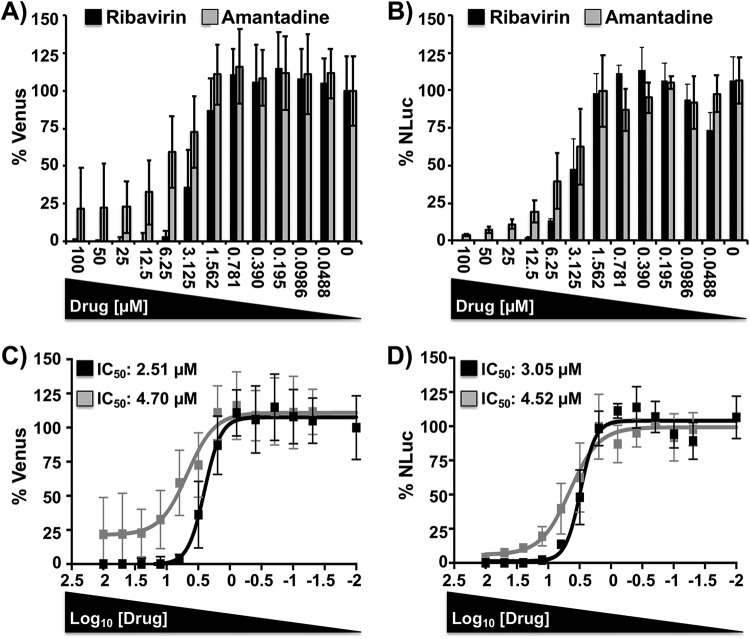
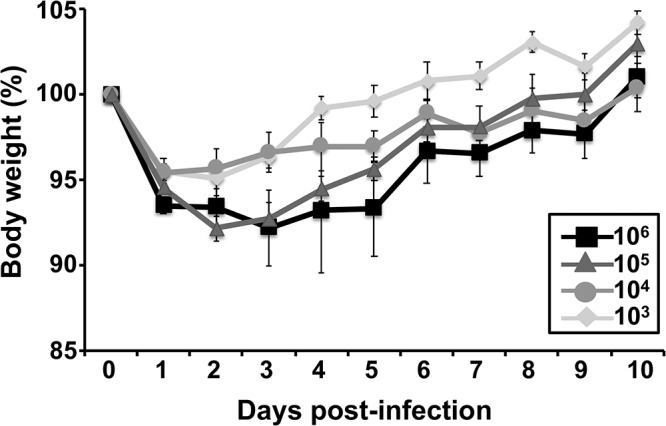
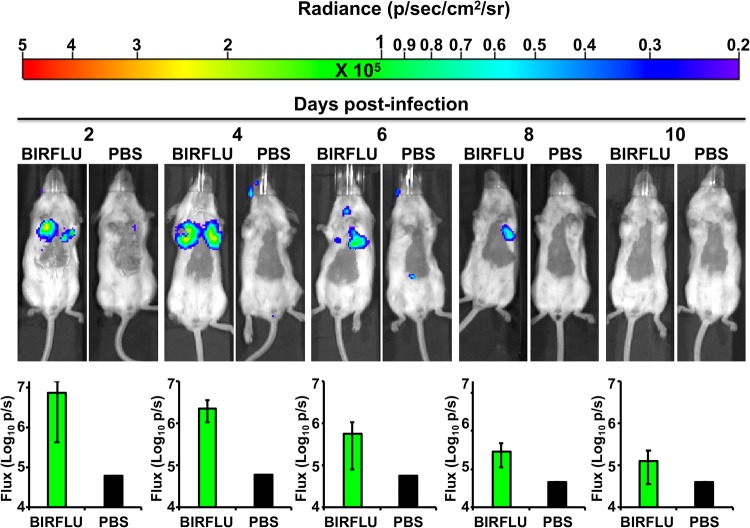
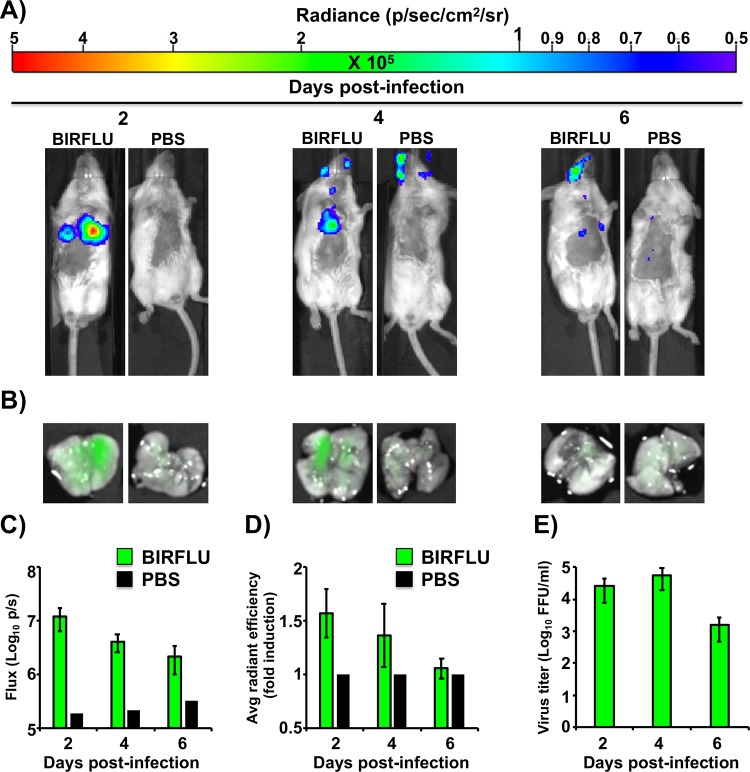
Studying influenza A virus (IAV) requires the use of secondary approaches to detect the presence of virus in infected cells. To overcome this problem, we and others have generated recombinant IAV expressing fluorescent or luciferase reporter genes. These foreign reporter genes can be used as valid surrogates to track the presence of virus. However, the limited capacity for incorporating foreign sequences in the viral genome forced researchers to select a fluorescent or a luciferase reporter gene, depending on the type of study. To circumvent this limitation, we engineered a novel recombinant replication-competent bireporter IAV (BIRFLU) expressing both fluorescent and luciferase reporter genes. In cultured cells, BIRFLU displayed growth kinetics comparable to those of wild-type (WT) virus and was used to screen neutralizing antibodies or compounds with antiviral activity. The expression of two reporter genes allows monitoring of viral inhibition by fluorescence or bioluminescence, overcoming the limitations associated with the use of one reporter gene as a readout. In vivo, BIRFLU effectively infected mice, and both reporter genes were detected using in vivo imaging systems (IVIS). The ability to generate recombinant IAV harboring multiple foreign genes opens unique possibilities for studying virus-host interactions and for using IAV in high-throughput screenings (HTS) to identify novel antivirals that can be incorporated into the therapeutic armamentarium to control IAV infections. Moreover, the ability to genetically manipulate the viral genome to express two foreign genes offers the possibility of developing novel influenza vaccines and the feasibility for using recombinant IAV as vaccine vectors to treat other pathogen infections.IMPORTANCE Influenza A virus (IAV) causes a human respiratory disease that is associated with significant health and economic consequences. In recent years, the use of replication-competent IAV expressing an easily traceable fluorescent or luciferase reporter protein has significantly contributed to progress in influenza research. However, researchers have been forced to select a fluorescent or a luciferase reporter gene due to the restricted capacity of the influenza viral genome for including foreign sequences. To overcome this limitation, we generated, for the first time, a recombinant replication-competent bireporter IAV (BIRFLU) that stably expresses two reporter genes (one fluorescent and one luciferase) to track IAV infections in vitro and in vivo The combination of cutting-edge techniques from molecular biology, animal research, and imaging technologies brings researchers the unique opportunity to use this new generation of reporter-expressing IAV to study viral infection dynamics in both cultured cells and animal models of viral infection.
Keywords: Nano luciferase; Venus fluorescence; bioluminescence; fluorescence; in vivo expression technology; influenza; reporter genes.
Copyright © 2019 American Society for Microbiology.
Figures
References
-
- Palese PM. 2007. Orthomyxoviridae: the viruses, and their replication In Knipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE (ed), Fields virology, 5th ed Lippincott Williams & Wilkins, Philadelphia, PA.
-
- Munster VJ, Baas C, Lexmond P, Waldenstrom J, Wallensten A, Fransson T, Rimmelzwaan GF, Beyer WE, Schutten M, Olsen B, Osterhaus AD, Fouchier RA. 2007. Spatial, temporal, and species variation in prevalence of influenza A viruses in wild migratory birds. PLoS Pathog 3:e61. doi: 10.1371/journal.ppat.0030061. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
