

Viral Factors Important for Efficient Replication of Influenza A Viruses in Cells of the Central Nervous System
- PMID: 30867311
- PMCID: PMC6532103
- DOI: 10.1128/JVI.02273-18
Viral Factors Important for Efficient Replication of Influenza A Viruses in Cells of the Central Nervous System
Abstract
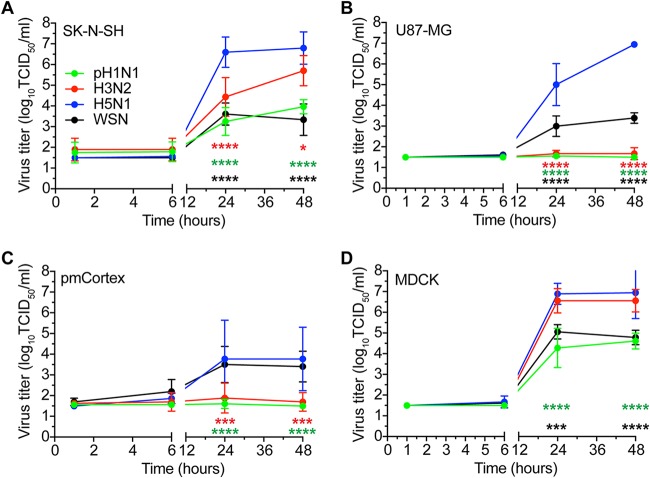
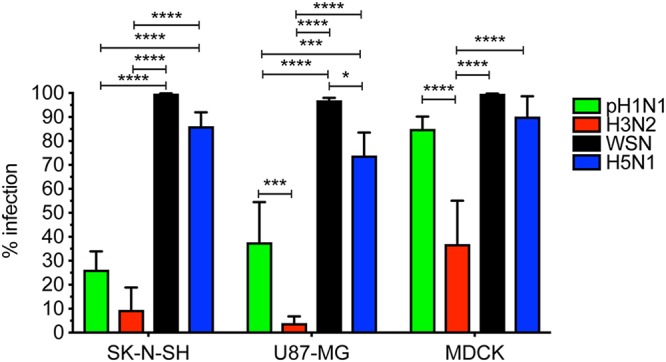
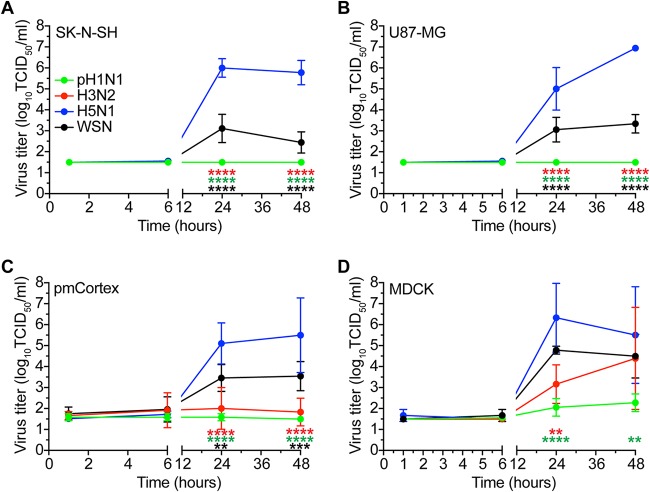
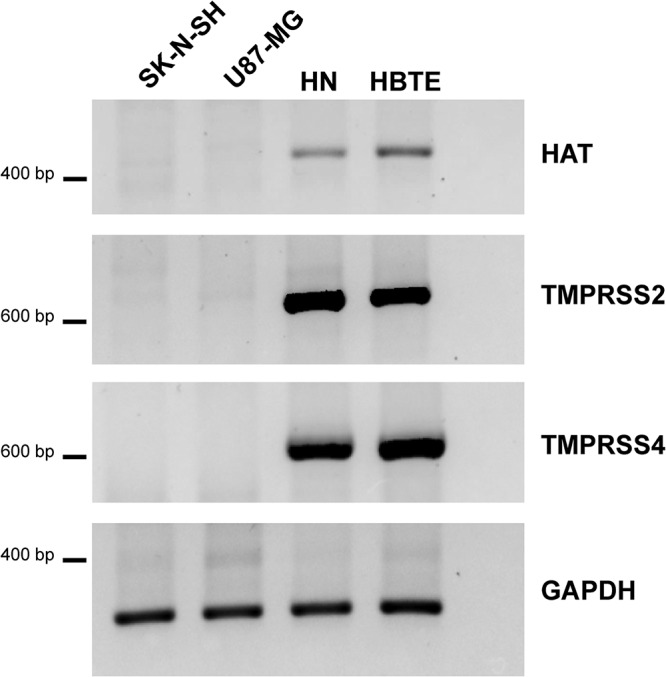
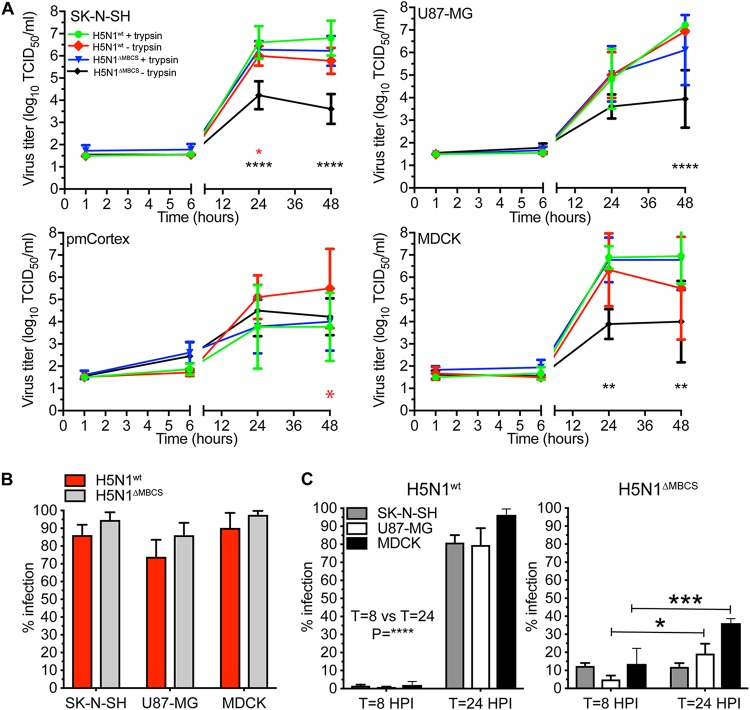
Central nervous system (CNS) disease is one of the most common extrarespiratory tract complications of influenza A virus infections. Remarkably, zoonotic H5N1 virus infections are more frequently associated with CNS disease than seasonal or pandemic influenza viruses. Little is known about the interaction between influenza A viruses and cells of the CNS; therefore, it is currently unknown which viral factors are important for efficient replication. Here, we determined the replication kinetics of a seasonal, pandemic, zoonotic, and lab-adapted influenza A virus in human neuron-like (SK-N-SH) and astrocyte-like (U87-MG) cells and primary mouse cortex neurons. In general, highly pathogenic avian influenza (HPAI) H5N1 virus replicated most efficiently in all cells, which was associated with efficient attachment and infection. Seasonal H3N2 and to a lesser extent pandemic H1N1 virus replicated in a trypsin-dependent manner in SK-N-SH but not in U87-MG cells. In the absence of trypsin, only HPAI H5N1 and WSN viruses replicated. Removal of the multibasic cleavage site (MBCS) from HPAI H5N1 virus attenuated, but did not abrogate, replication. Taken together, our results showed that the MBCS and, to a lesser extent, the ability to attach are important determinants for efficient replication of HPAI H5N1 virus in cells of the CNS. This suggests that both an alternative hemagglutinin (HA) cleavage mechanism and preference for α-2,3-linked sialic acids allowing efficient attachment contribute to the ability of influenza A viruses to replicate efficiently in cells of the CNS. This study further improves our knowledge on potential viral factors important for the neurotropic potential of influenza A viruses.IMPORTANCE Central nervous system (CNS) disease is one of the most common extrarespiratory tract complications of influenza A virus infections, and the frequency and severity differ between seasonal, pandemic, and zoonotic influenza viruses. However, little is known about the interaction of these viruses with cells of the CNS. Differences among seasonal, pandemic, and zoonotic influenza viruses in replication efficacy in CNS cells, in vitro, suggest that the presence of an alternative HA cleavage mechanism and ability to attach are important viral factors. Identifying these viral factors and detailed knowledge of the interaction between influenza virus and CNS cells are important to prevent and treat this potentially lethal CNS disease.
Keywords: CNS disease; H1N1; H3N2; H5N1; encephalitis; extrarespiratory; influenza A virus; pathogenesis; viral replication; virus attachment.
Copyright © 2019 Siegers et al.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
