

Conditional knockout of UBC13 produces disturbances in gait and spontaneous locomotion and exploration in mice
- PMID: 30867488
- PMCID: PMC6416404
- DOI: 10.1038/s41598-019-40714-3
Conditional knockout of UBC13 produces disturbances in gait and spontaneous locomotion and exploration in mice
Abstract
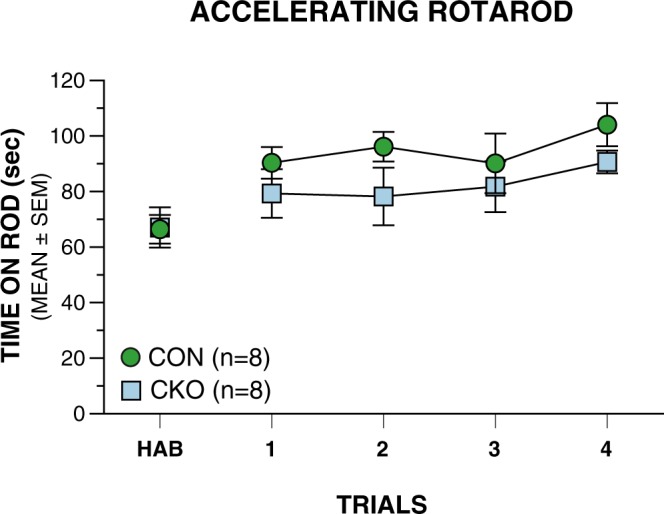
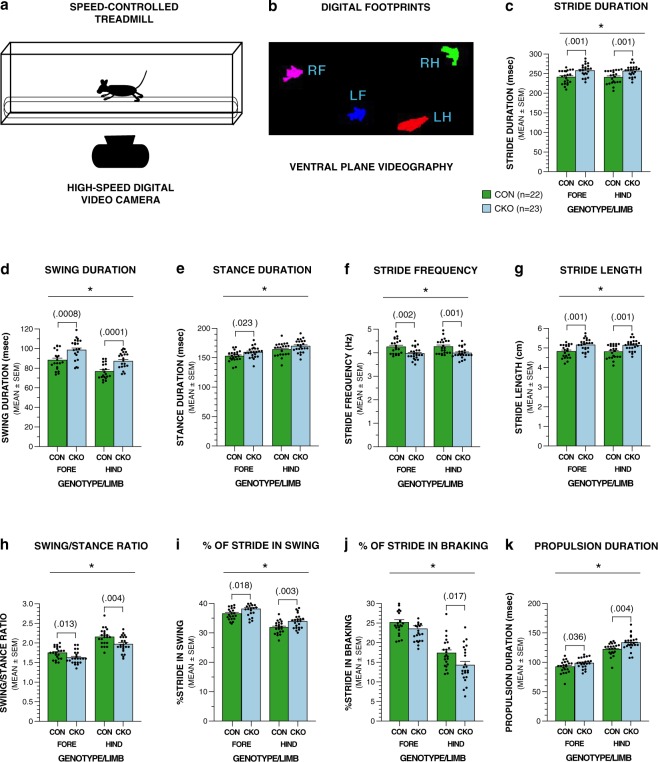
Here we have characterized the functional impairments resulting from conditional knockout of the ubiquitin-conjugating E2 enzyme (UBC13) in rodent cerebellar granule neurons, which greatly increases the parallel fiber presynaptic boutons and functional parallel fiber/Purkinje cell synapses. We report that conditional UBC13 knockout mice exhibit reliable deficits on several gait-related variables when their velocity of ambulation is tightly controlled by a moving treadmill and by restricting space for movement. Selected gait parameters and movement patterns related to spontaneous exploration in an open field may also be affected in conditional UBC13 knockout mice. Analysis of open-field data as a function of test session half using force-plate actometer instrumentation suggest that conditional UBC13 knockout mice have alterations in emotionality, possibly affecting gait and movement variables. These findings suggest that conditional UBC13 knockout mice represent a valuable platform for assessing the effects of disturbances in cerebellar granule cell circuitry on gait and other aspects of locomotion. Also, the possibility that psychological factors such as altered emotionality may impact gait and movement patterns in these mice suggest that these mice may provide a useful model for evaluating analogous behavioral impairments in autism spectrum disorders and other neurodevelopmental syndromes associated with deregulation of ubiquitin signaling.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Altman, J. & Bayer, S. A. Development of the cerebellar system: in relation to its evolution, structure, and functions. pp. 783 (CRC Press, Boca Raton, 1997).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
