

Increased intron retention is a post-transcriptional signature associated with progressive aging and Alzheimer's disease
- PMID: 30868713
- PMCID: PMC6516162
- DOI: 10.1111/acel.12928
Increased intron retention is a post-transcriptional signature associated with progressive aging and Alzheimer's disease
Abstract
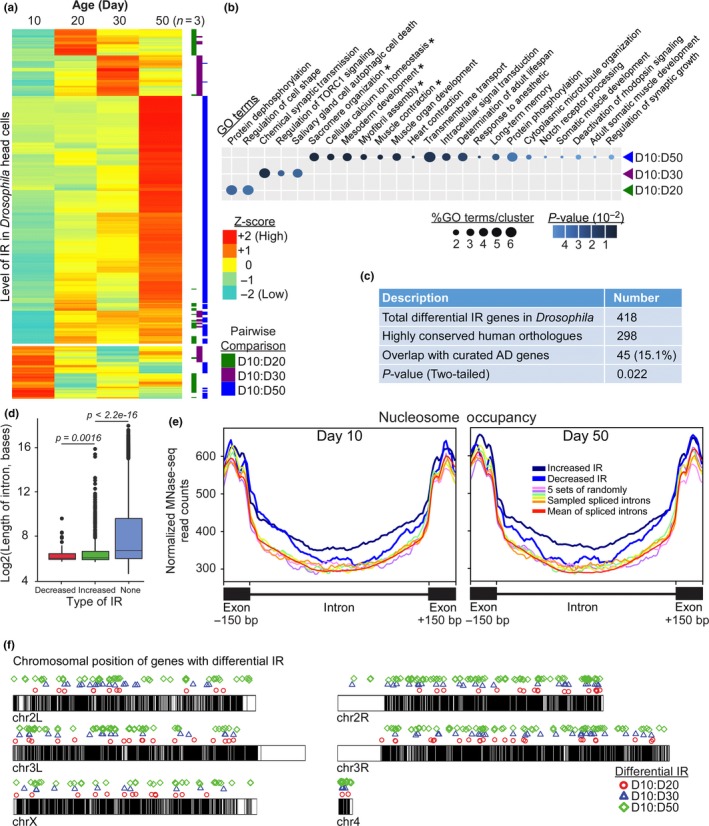
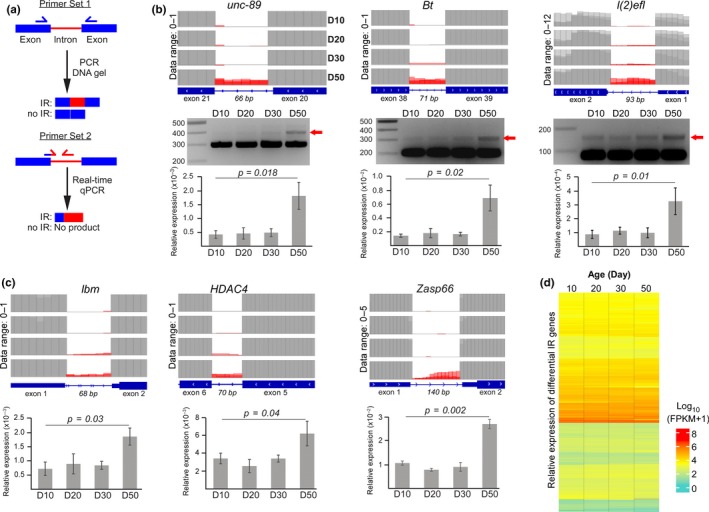
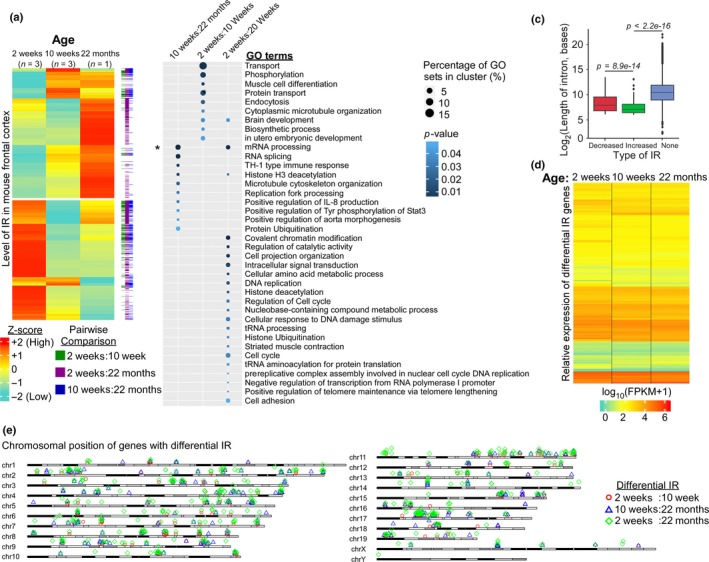
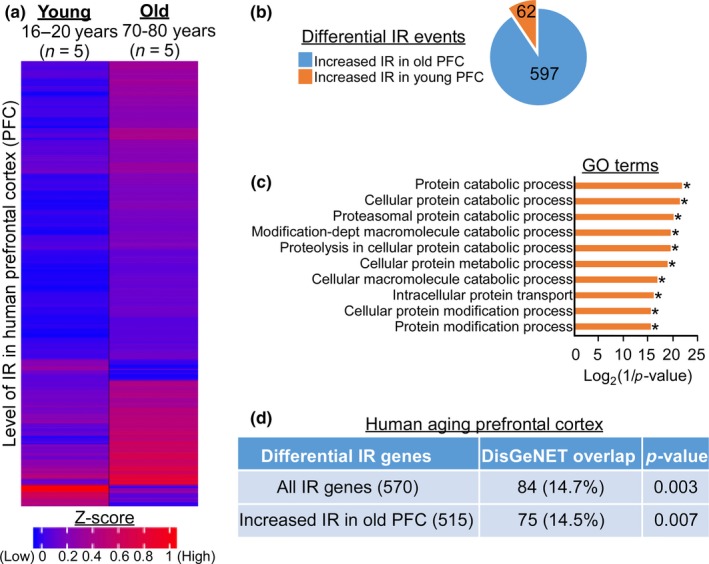
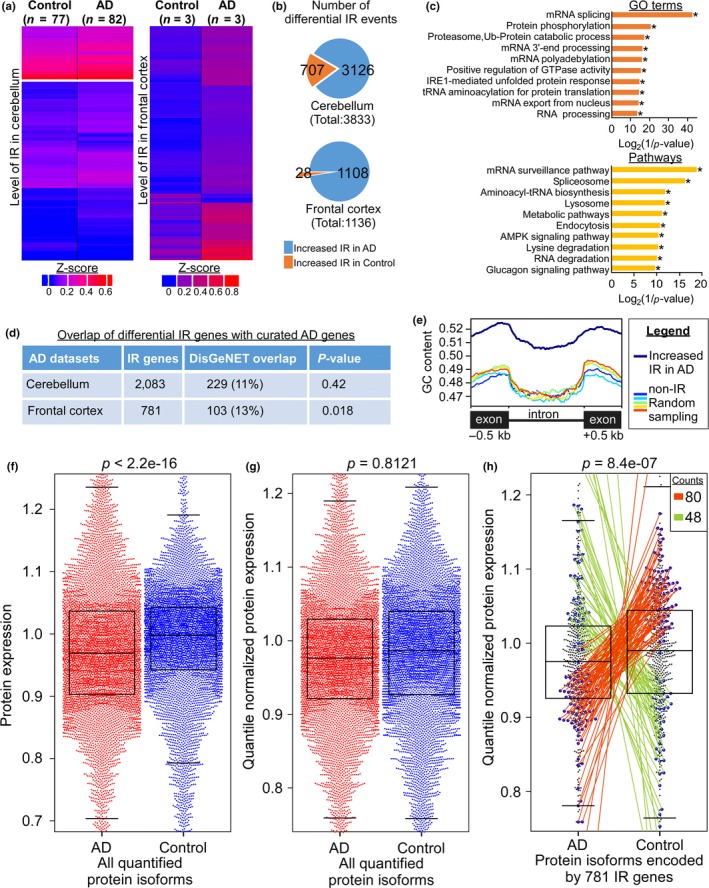
Intron retention (IR) by alternative splicing is a conserved regulatory mechanism that can affect gene expression and protein function during adult development and age-onset diseases. However, it remains unclear whether IR undergoes spatial or temporal changes during different stages of aging or neurodegeneration like Alzheimer's disease (AD). By profiling the transcriptome of Drosophila head cells at different ages, we observed a significant increase in IR events for many genes during aging. Differential IR affects distinct biological functions at different ages and occurs at several AD-associated genes in older adults. The increased nucleosome occupancy at the differentially retained introns in young animals suggests that it may regulate the level of IR during aging. Notably, an increase in the number of IR events was also observed in healthy older mouse and human brain tissues, as well as in the cerebellum and frontal cortex from independent AD cohorts. Genes with differential IR shared many common features, including shorter intron length, no perturbation in their mRNA level, and enrichment for biological functions that are associated with mRNA processing and proteostasis. The differentially retained introns identified in AD frontal cortex have higher GC content, with many of their mRNA transcripts showing an altered level of protein expression compared to control samples. Taken together, our results suggest that an increased IR is an conserved signature that is associated with aging. By affecting pathways involved in mRNA and protein homeostasis, changes of IR pattern during aging may regulate the transition from healthy to pathological state in late-onset sporadic AD.
Keywords: Alzheimer’s disease; Drosophila; Intron retention; aging; human; mouse.
© 2019 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
None declared.
Figures
References
-
- Audic, S. , & Claverie, J. M. (1997). The significance of digital gene expression profiles. Genome Research, 7, 986–995. - PubMed
-
- Bai, B. , Hales, C. M. , Chen, P.‐C. , Gozal, Y. , Dammer, E. B. , Fritz, J. J. , … Peng, J. (2013). U1 small nuclear ribonucleoprotein complex and RNA splicing alterations in Alzheimer’s disease. Proceedings of the National Academy of Sciences, 110, 16562–16567. 10.1073/pnas.1310249110 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
