

The small non-coding RNA RsaE influences extracellular matrix composition in Staphylococcus epidermidis biofilm communities
- PMID: 30870530
- PMCID: PMC6435200
- DOI: 10.1371/journal.ppat.1007618
The small non-coding RNA RsaE influences extracellular matrix composition in Staphylococcus epidermidis biofilm communities
Abstract
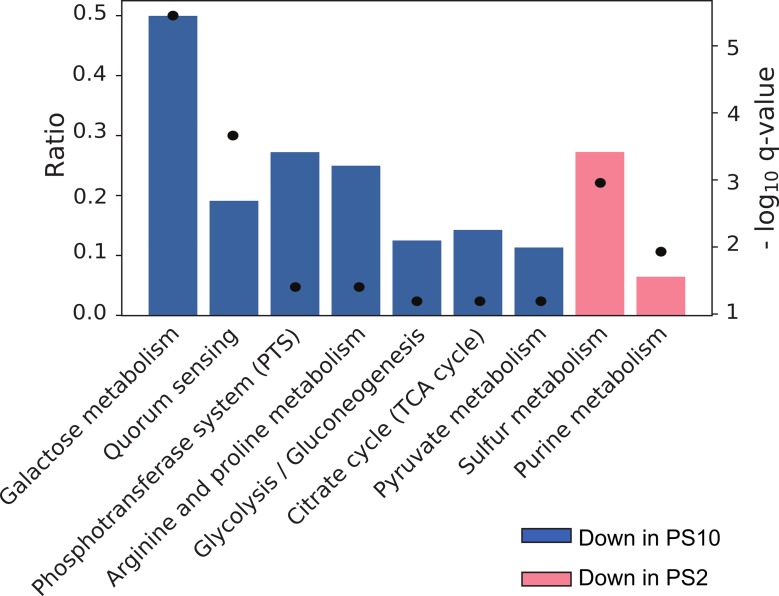
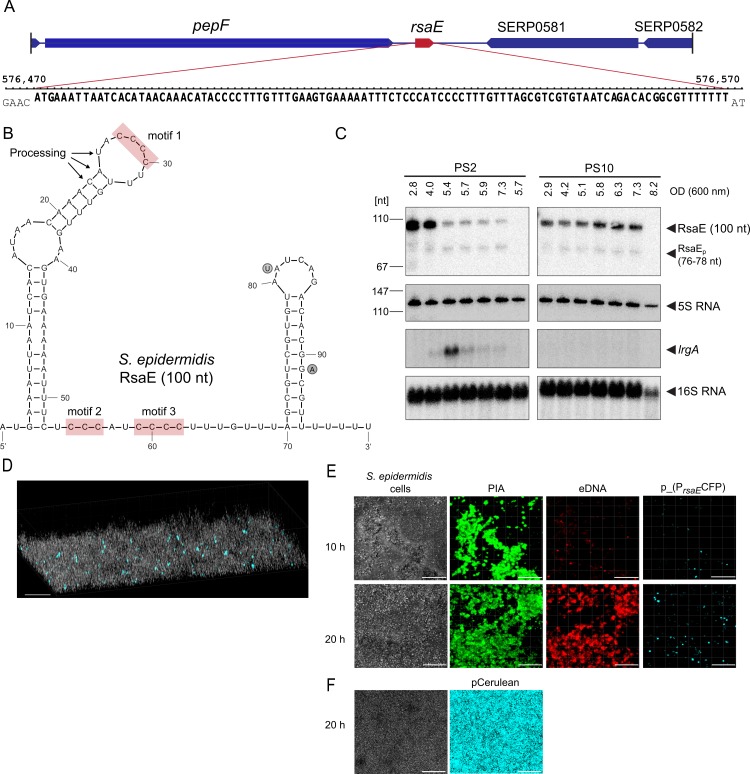
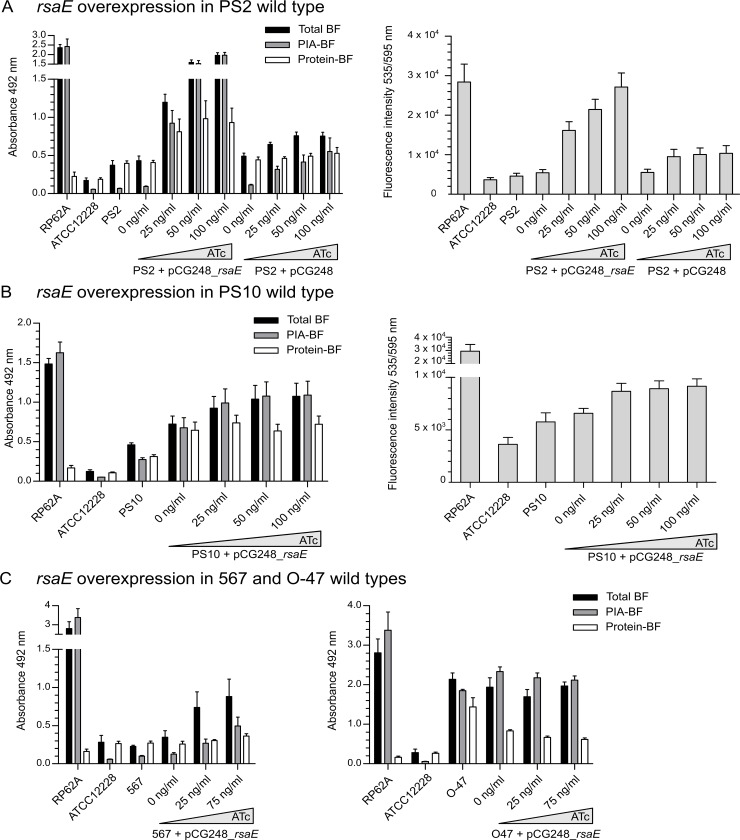
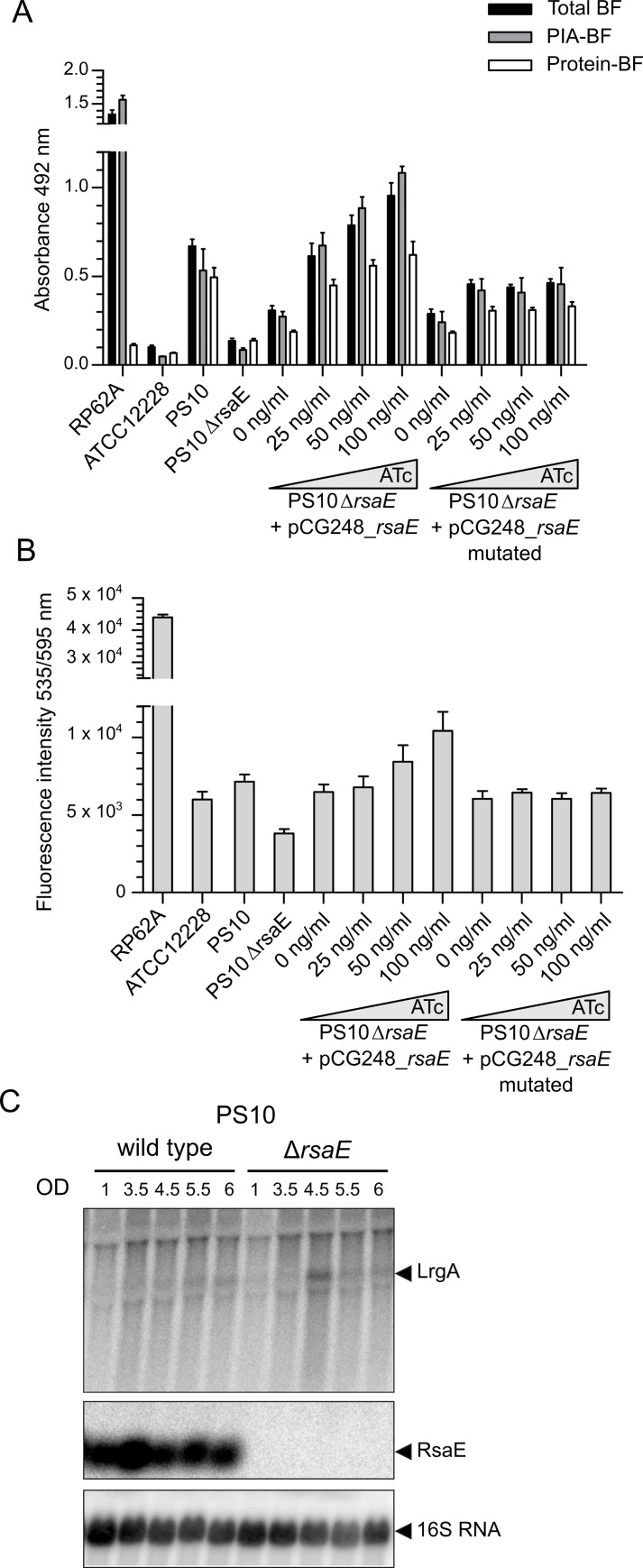
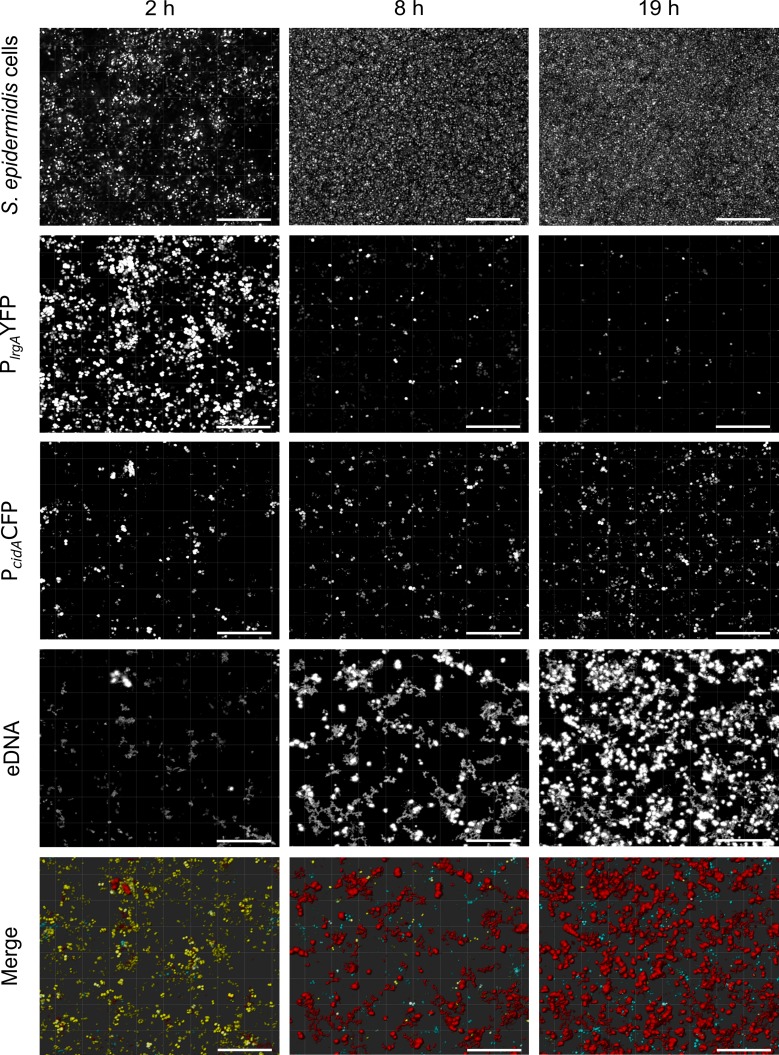
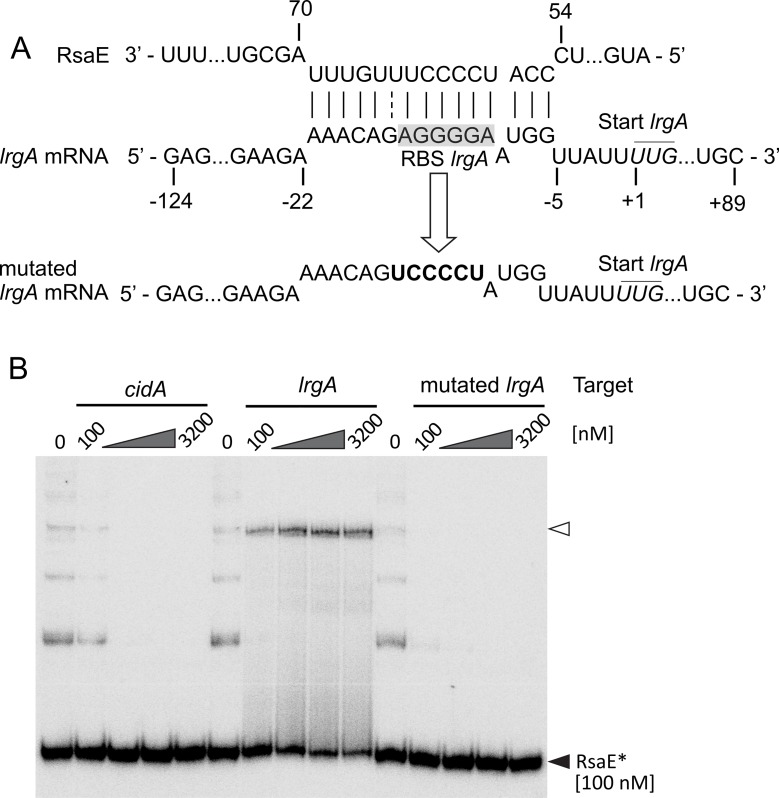
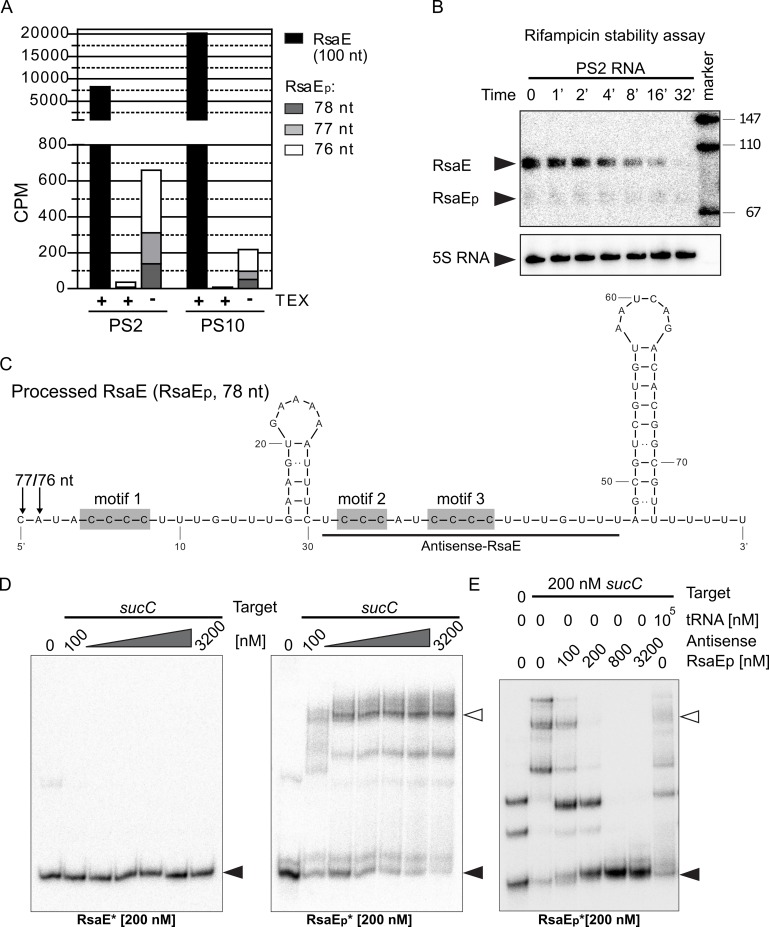
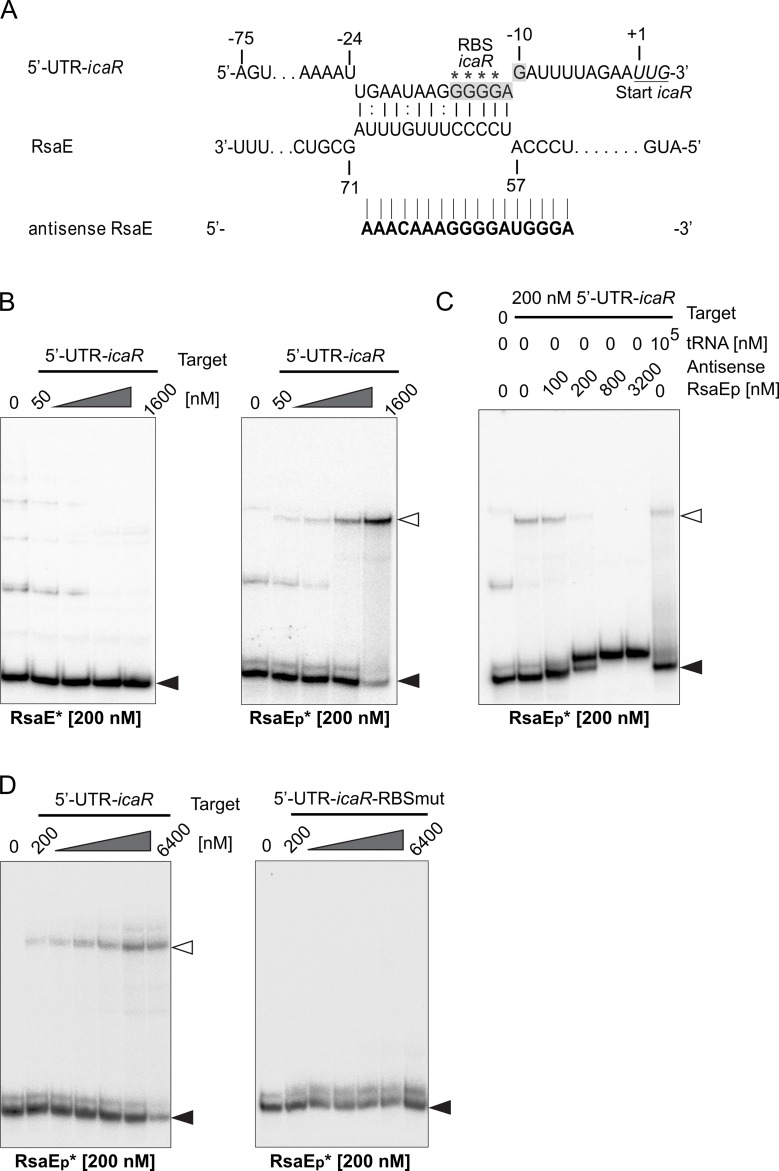
RsaE is a conserved small regulatory RNA (sRNA) which was previously reported to represent a riboregulator of central carbon flow and other metabolic pathways in Staphylococcus aureus and Bacillus subtilis. Here we show that RsaE contributes to extracellular (e)DNA release and biofilm-matrix switching towards polysaccharide intercellular adhesin (PIA) production in a hypervariable Staphylococcus epidermidis isolate. Transcriptome analysis through differential RNA sequencing (dRNA-seq) in combination with confocal laser scanning microscopy (CLSM) and reporter gene fusions demonstrate that S. epidermidis protein- and PIA-biofilm matrix producers differ with respect to RsaE and metabolic gene expression. RsaE is spatiotemporally expressed within S. epidermidis PIA-mediated biofilms, and its overexpression triggers a PIA biofilm phenotype as well as eDNA release in an S. epidermidis protein biofilm matrix-producing strain background. dRNA-seq and Northern blot analyses revealed RsaE to exist as a major full-length 100-nt transcript and a minor processed species lacking approximately 20 nucleotides at the 5'-end. RsaE processing results in expansion of the mRNA target spectrum. Thus, full-length RsaE interacts with S. epidermidis antiholin-encoding lrgA mRNA, facilitating bacterial lysis and eDNA release. Processed RsaE, however, interacts with the 5'-UTR of icaR and sucCD mRNAs, encoding the icaADBC biofilm operon repressor IcaR and succinyl-CoA synthetase of the tricarboxylic acid (TCA) cycle, respectively. RsaE augments PIA-mediated biofilm matrix production, most likely through activation of icaADBC operon expression via repression of icaR as well as by TCA cycle inhibition and re-programming of staphylococcal central carbon metabolism towards PIA precursor synthesis. Additionally, RsaE supports biofilm formation by mediating the release of eDNA as stabilizing biofilm matrix component. As RsaE itself is heterogeneously expressed within biofilms, we consider this sRNA to function as a factor favoring phenotypic heterogeneity and supporting division of labor in S. epidermidis biofilm communities.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
