

The Impact of the C-Terminal Region on the Interaction of Topoisomerase II Alpha with Mitotic Chromatin
- PMID: 30871006
- PMCID: PMC6429393
- DOI: 10.3390/ijms20051238
The Impact of the C-Terminal Region on the Interaction of Topoisomerase II Alpha with Mitotic Chromatin
Abstract
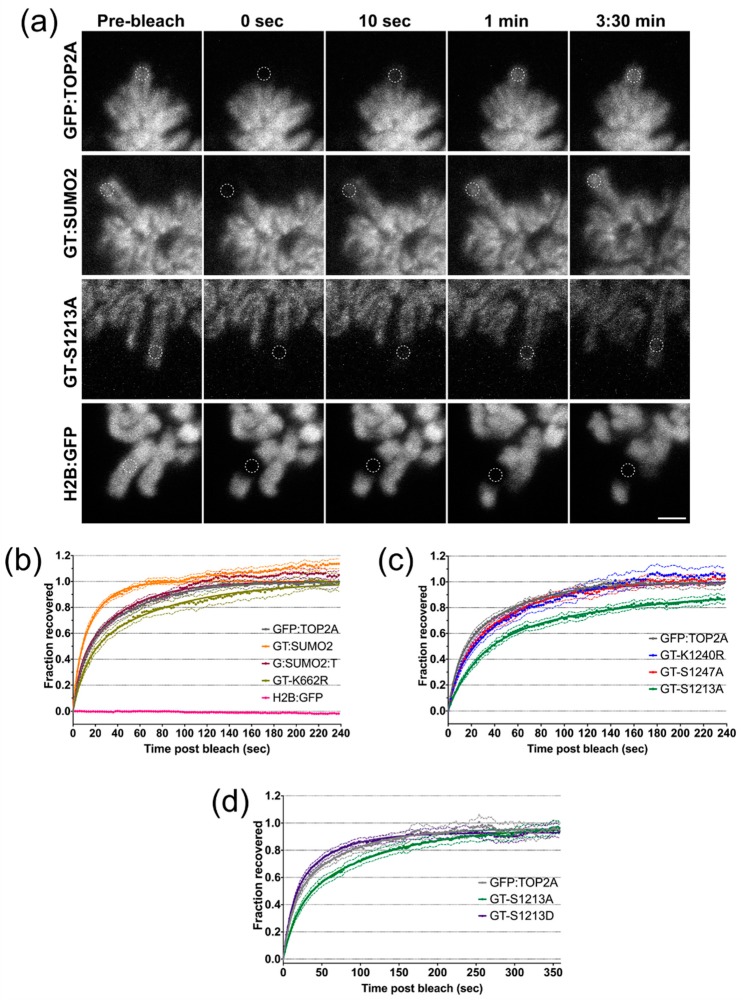
Type II topoisomerase enzymes are essential for resolving DNA topology problems arising through various aspects of DNA metabolism. In vertebrates two isoforms are present, one of which (TOP2A) accumulates on chromatin during mitosis. Moreover, TOP2A targets the mitotic centromere during prophase, persisting there until anaphase onset. It is the catalytically-dispensable C-terminal domain of TOP2 that is crucial in determining this isoform-specific behaviour. In this study we show that, in addition to the recently identified chromatin tether domain, several other features of the alpha-C-Terminal Domain (CTD). influence the mitotic localisation of TOP2A. Lysine 1240 is a major SUMOylation target in cycling human cells and the efficiency of this modification appears to be influenced by T1244 and S1247 phosphorylation. Replacement of K1240 by arginine results in fewer cells displaying centromeric TOP2A accumulation during prometaphase-metaphase. The same phenotype is displayed by cells expressing TOP2A in which either of the mitotic phosphorylation sites S1213 or S1247 has been substituted by alanine. Conversely, constitutive modification of TOP2A by fusion to SUMO2 exerts the opposite effect. FRAP analysis of protein mobility indicates that post-translational modification of TOP2A can influence the enzyme's residence time on mitotic chromatin, as well as its subcellular localisation.
Keywords: C-terminal domain; SUMO; centromere; chromosome; mitosis; phosphorylation; post-translational modification; topoisomerase II alpha.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
