

Secreted PD-L1 variants mediate resistance to PD-L1 blockade therapy in non-small cell lung cancer
- PMID: 30872362
- PMCID: PMC6446862
- DOI: 10.1084/jem.20180870
Secreted PD-L1 variants mediate resistance to PD-L1 blockade therapy in non-small cell lung cancer
Abstract
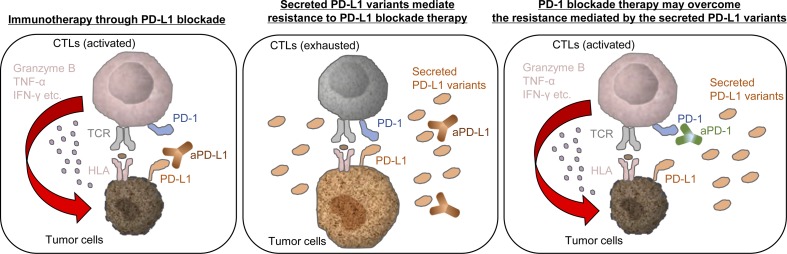
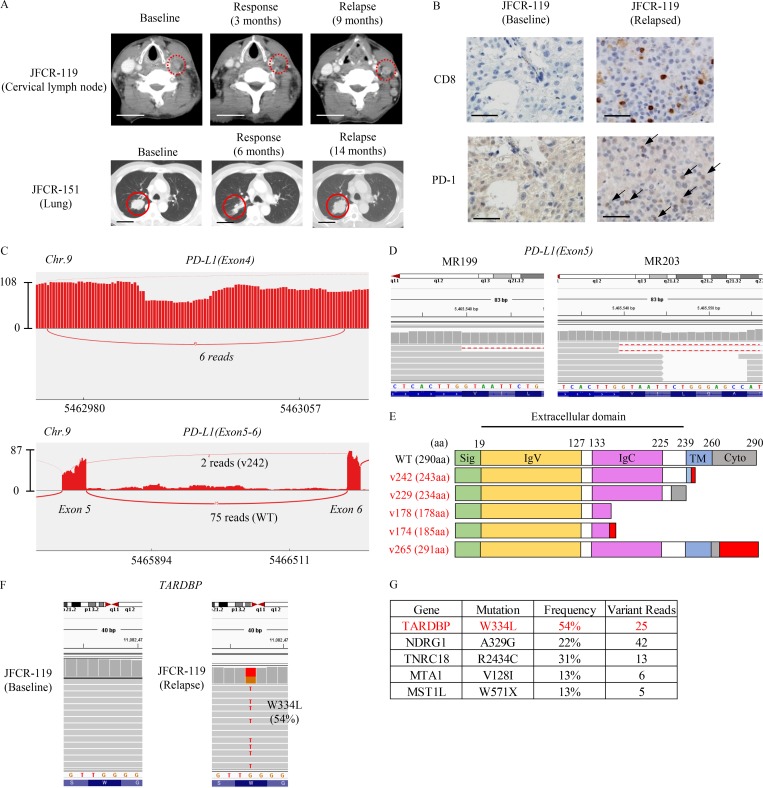
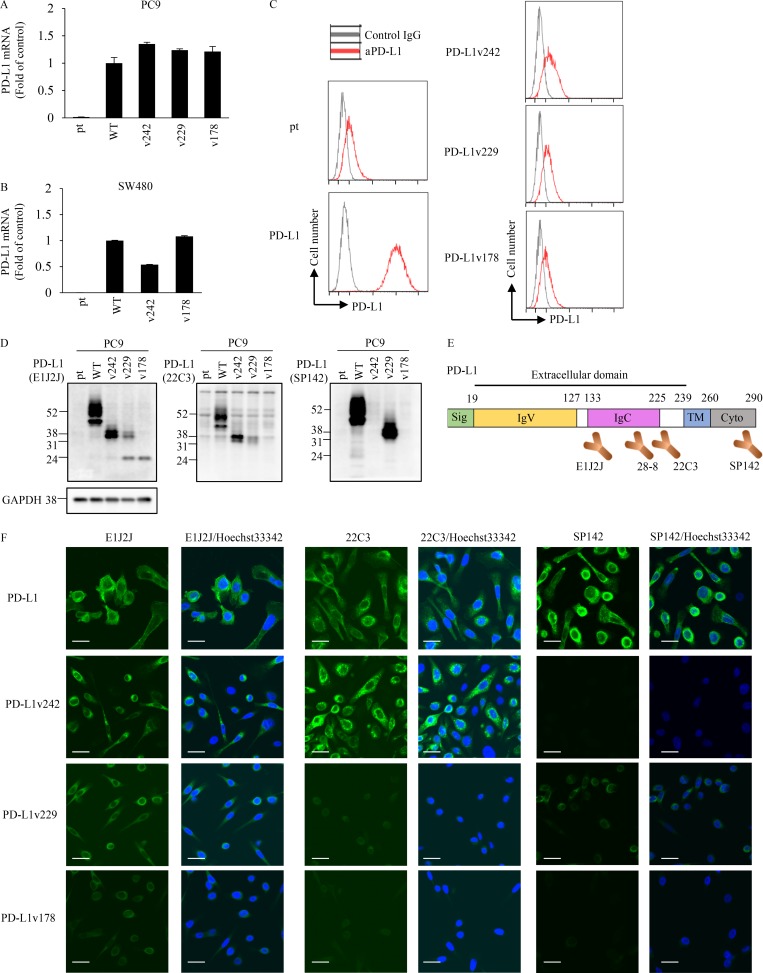
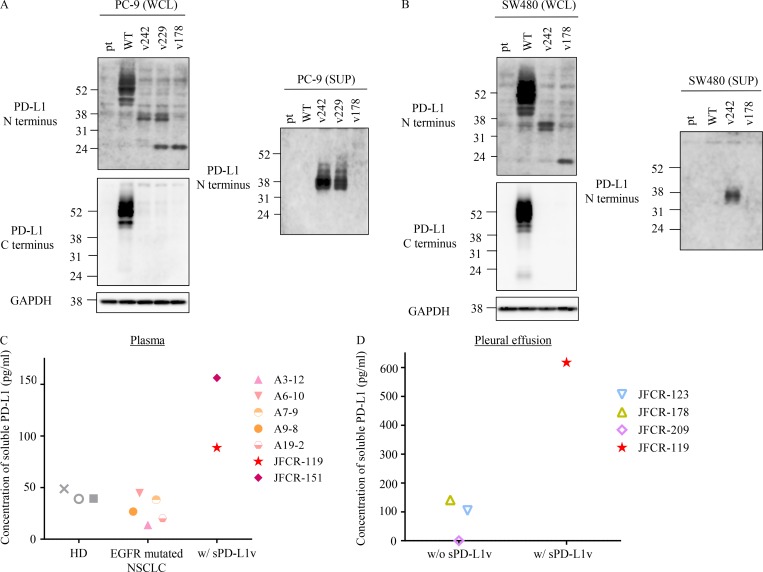
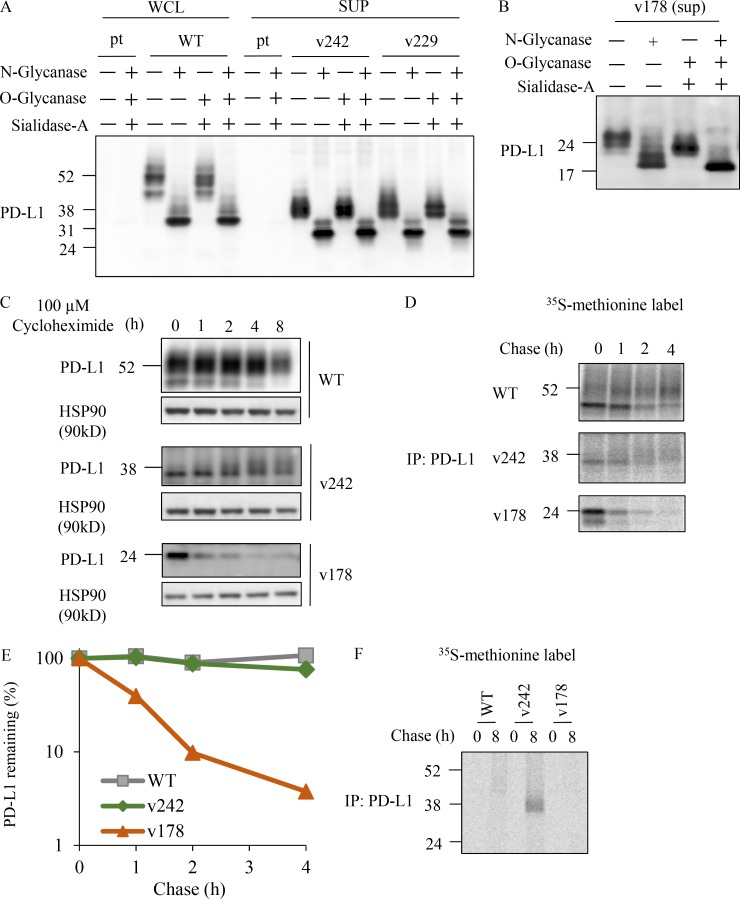
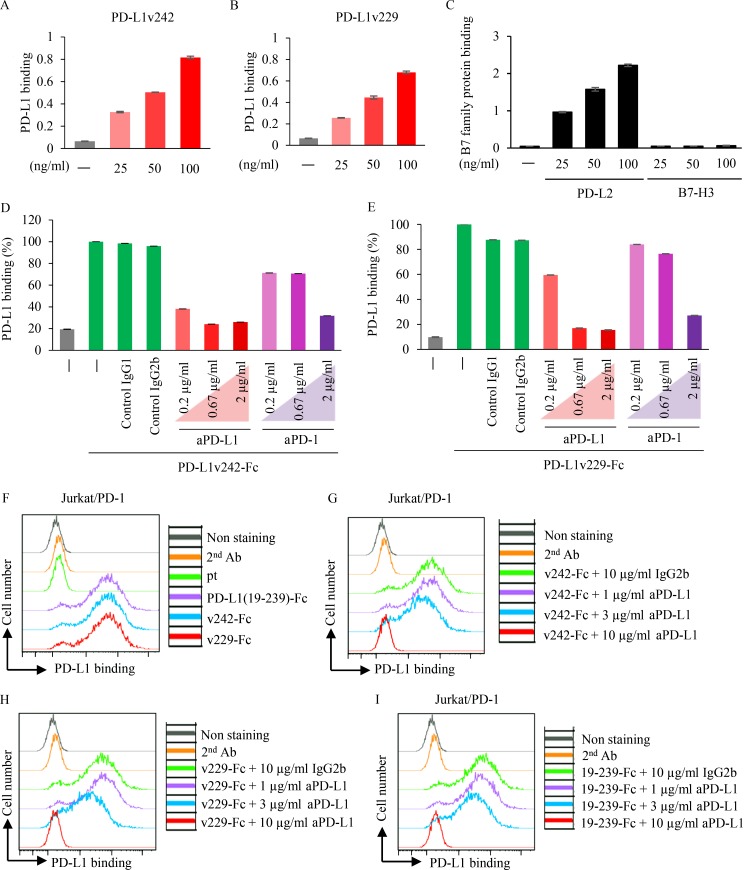
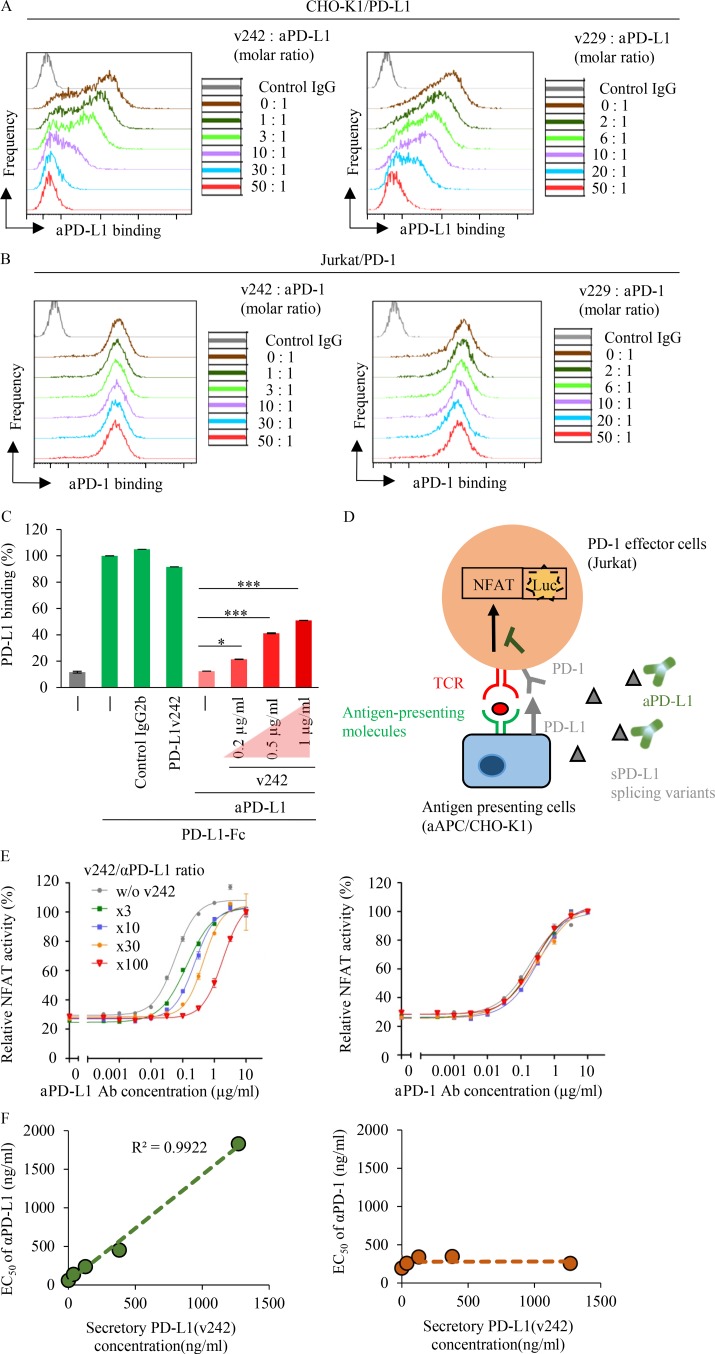
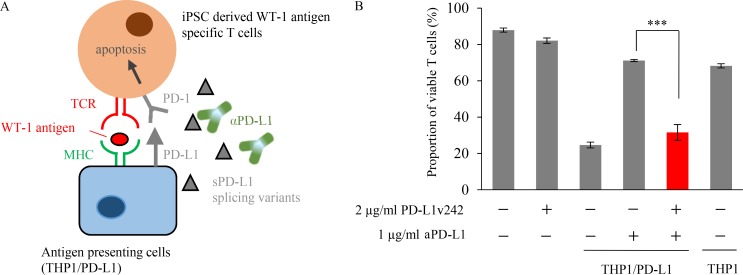
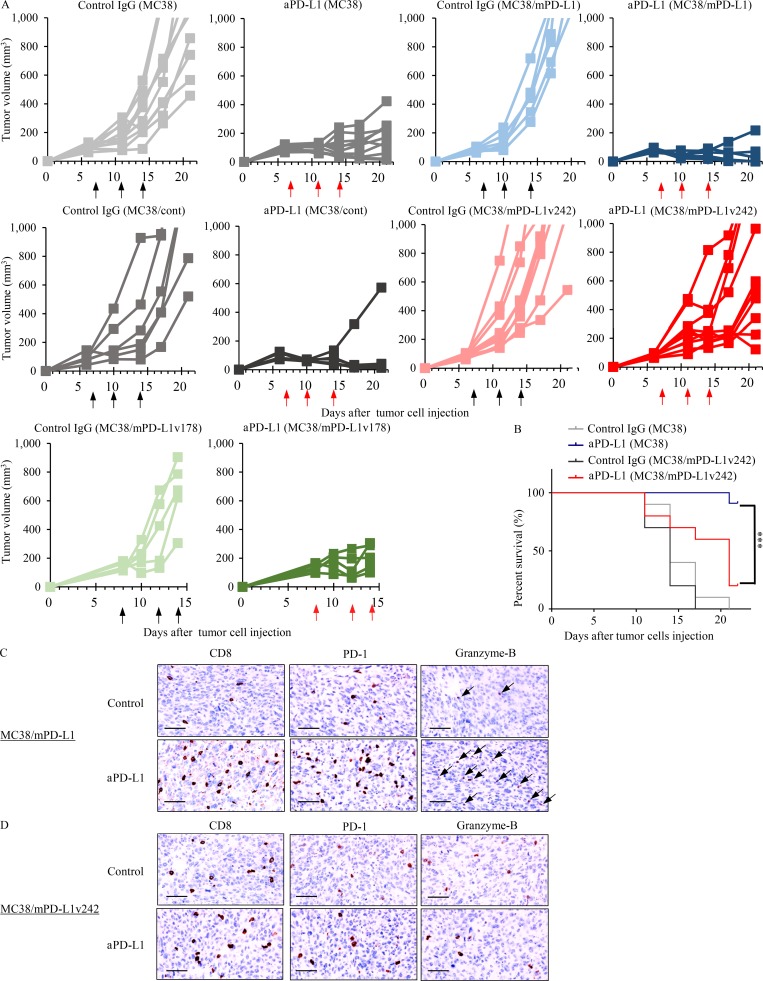
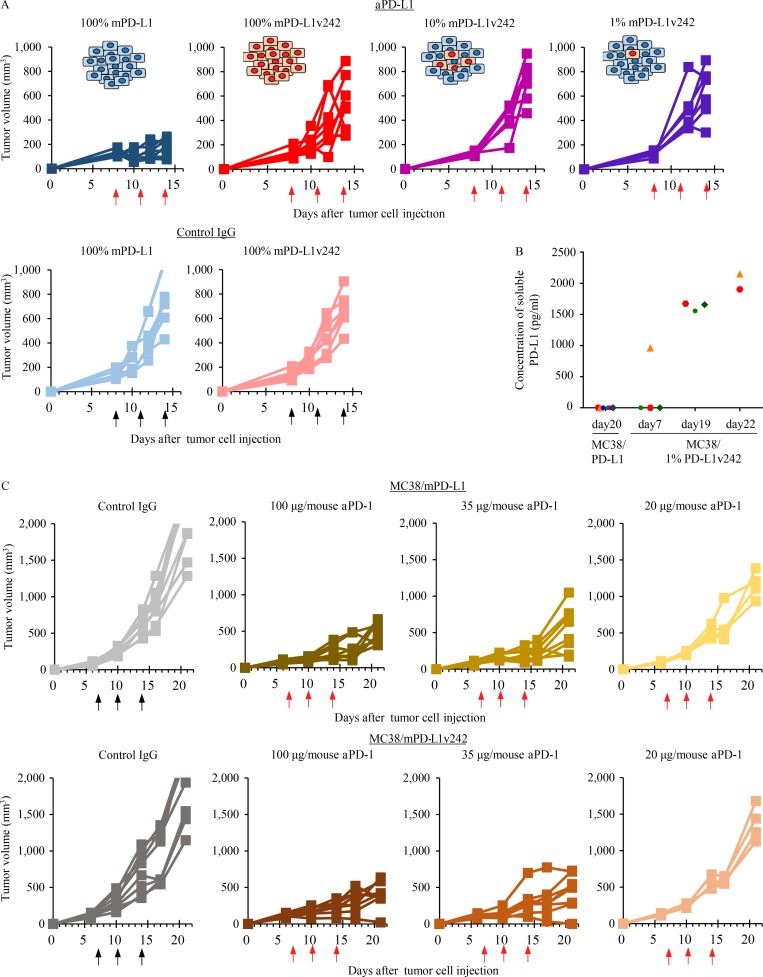
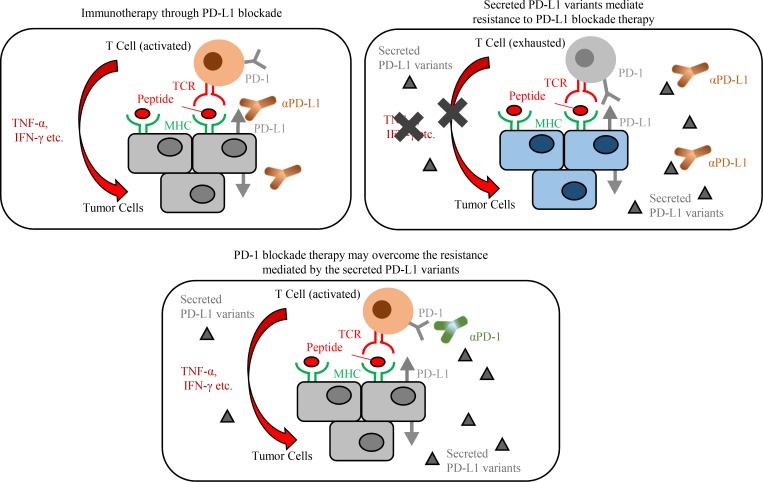
Immune checkpoint blockade against programmed cell death 1 (PD-1) and its ligand PD-L1 often induces durable tumor responses in various cancers, including non-small cell lung cancer (NSCLC). However, therapeutic resistance is increasingly observed, and the mechanisms underlying anti-PD-L1 (aPD-L1) antibody treatment have not been clarified yet. Here, we identified two unique secreted PD-L1 splicing variants, which lacked the transmembrane domain, from aPD-L1-resistant NSCLC patients. These secreted PD-L1 variants worked as "decoys" of aPD-L1 antibody in the HLA-matched coculture system of iPSC-derived CD8 T cells and cancer cells. Importantly, mixing only 1% MC38 cells with secreted PD-L1 variants and 99% of cells that expressed wild-type PD-L1 induced resistance to PD-L1 blockade in the MC38 syngeneic xenograft model. Moreover, anti-PD-1 (aPD-1) antibody treatment overcame the resistance mediated by the secreted PD-L1 variants. Collectively, our results elucidated a novel resistant mechanism of PD-L1 blockade antibody mediated by secreted PD-L1 variants.
© 2019 Gong et al.
Figures
References
-
- Arnold E.S., Ling S.C., Huelga S.C., Lagier-Tourenne C., Polymenidou M., Ditsworth D., Kordasiewicz H.B., McAlonis-Downes M., Platoshyn O., Parone P.A., et al. 2013. ALS-linked TDP-43 mutations produce aberrant RNA splicing and adult-onset motor neuron disease without aggregation or loss of nuclear TDP-43. Proc. Natl. Acad. Sci. USA. 110:E736–E745. 10.1073/pnas.1222809110 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
