

Persistence of Candida albicans in the Oral Mucosa Induces a Curbed Inflammatory Host Response That Is Independent of Immunosuppression
- PMID: 30873177
- PMCID: PMC6400982
- DOI: 10.3389/fimmu.2019.00330
Persistence of Candida albicans in the Oral Mucosa Induces a Curbed Inflammatory Host Response That Is Independent of Immunosuppression
Abstract
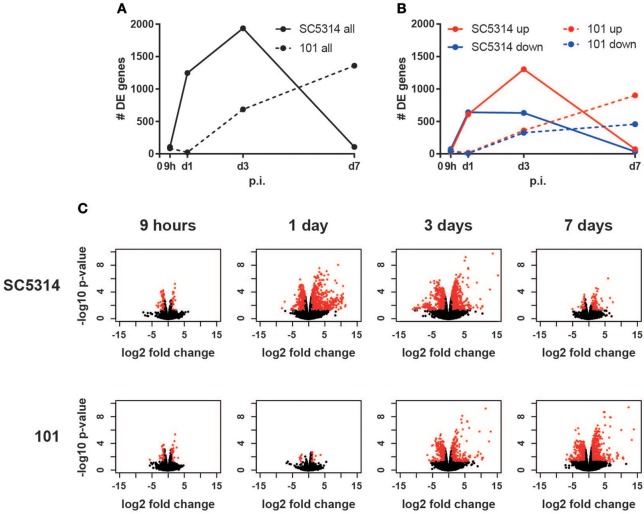
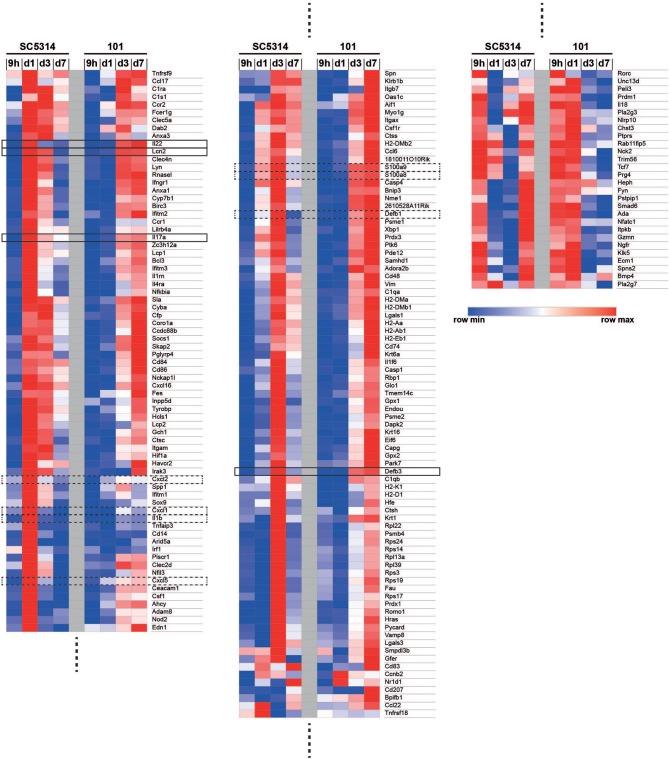
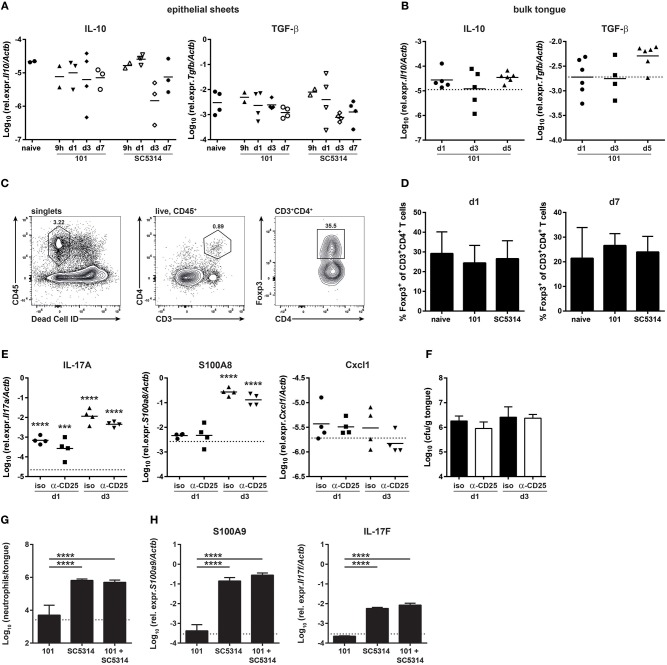
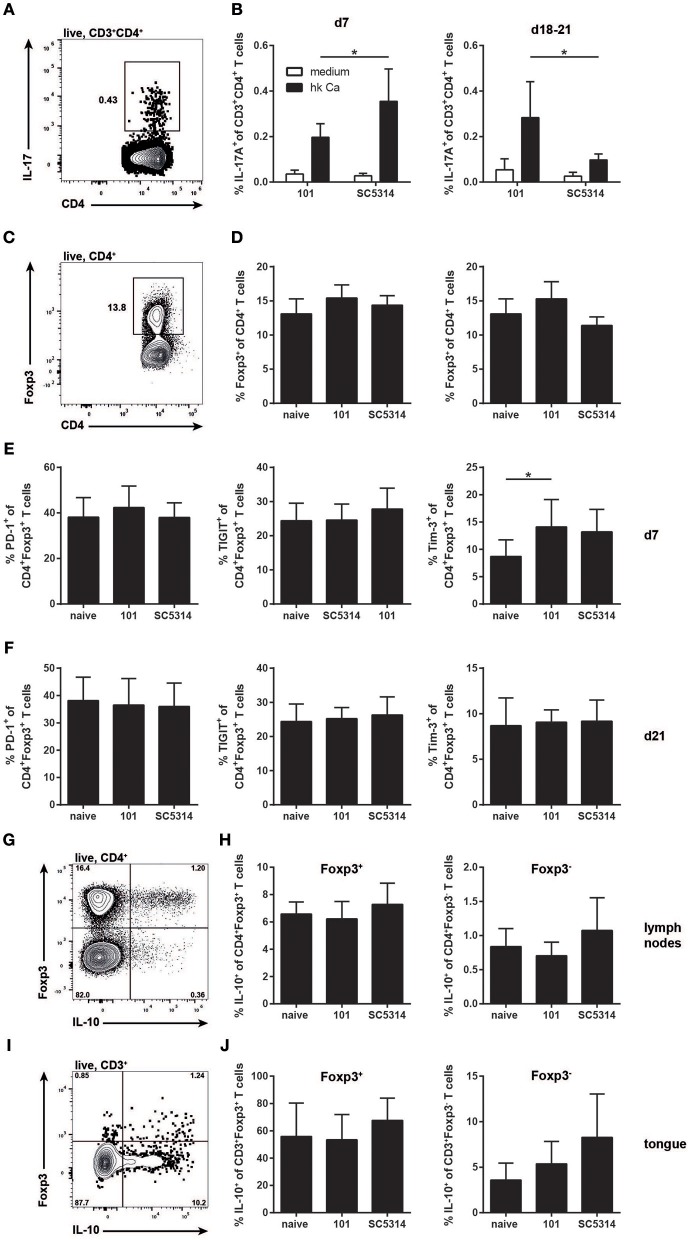
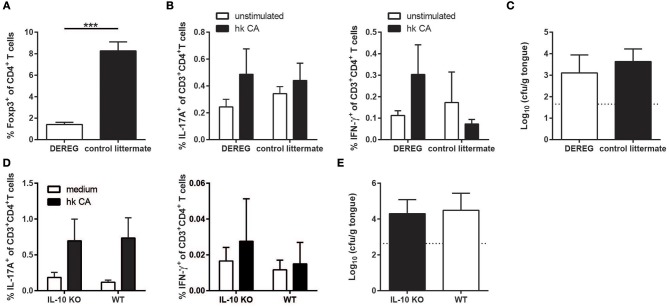
Controlled immune activation in response to commensal microbes is critical for the maintenance of stable colonization and prevention of microbial overgrowth on epithelial surfaces. Our understanding of the host mechanisms that regulate bacterial commensalism has increased substantially, however, much less data exist regarding host responses to members of the fungal microbiota on colonized surfaces. Using a murine model of oropharyngeal candidiasis, we have recently shown that differences in immune activation in response to diverse natural isolates of Candida albicans are associated with different outcomes of the host-fungal interaction. Here we applied a genome-wide transcriptomic approach to show that rapid induction of a strong inflammatory response characterized by neutrophil-associated genes upon C. albicans colonization inversely correlated with the ability of the fungus to persist in the oral mucosa. Surprisingly, persistent fungal isolates showed no signs of a compensatory regulatory immune response. By combining RNA-seq data, genetic mouse models, and co-infection experiments, we show that attenuation of the inflammatory response at the onset of infection with a persistent isolate is not a consequence of enhanced immunosuppression. Importantly, depletion of regulatory T cells or deletion of the immunoregulatory cytokine IL-10 did not alter host-protective type 17 immunity nor did it impair fungal survival in the oral mucosa, indicating that persistence of C. albicans in the oral mucosa is not a consequence of suppressed antifungal immunity.
Keywords: Candida albicans; IL-10; IL-17; immune regulation; oropharyngeal candidiasis; persistence; regulatory T cells.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases