

In vitro metabolic activation of vitamin D3 by using a multi-compartment microfluidic liver-kidney organ on chip platform
- PMID: 30874583
- PMCID: PMC6420623
- DOI: 10.1038/s41598-019-40851-9
In vitro metabolic activation of vitamin D3 by using a multi-compartment microfluidic liver-kidney organ on chip platform
Abstract
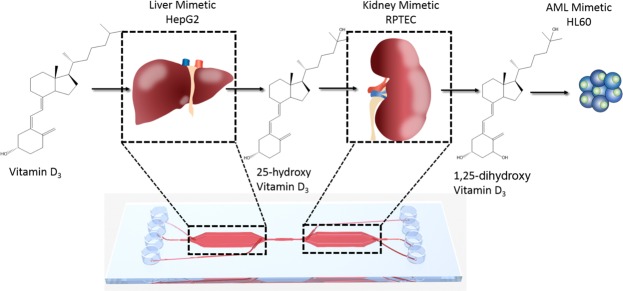
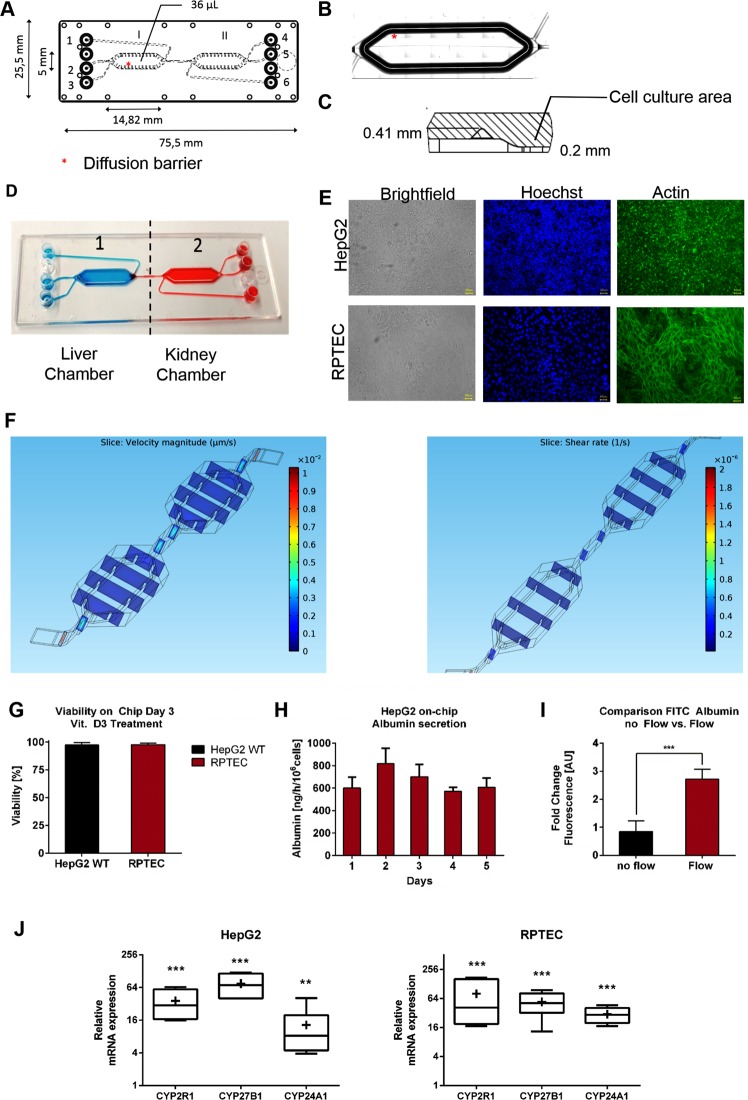
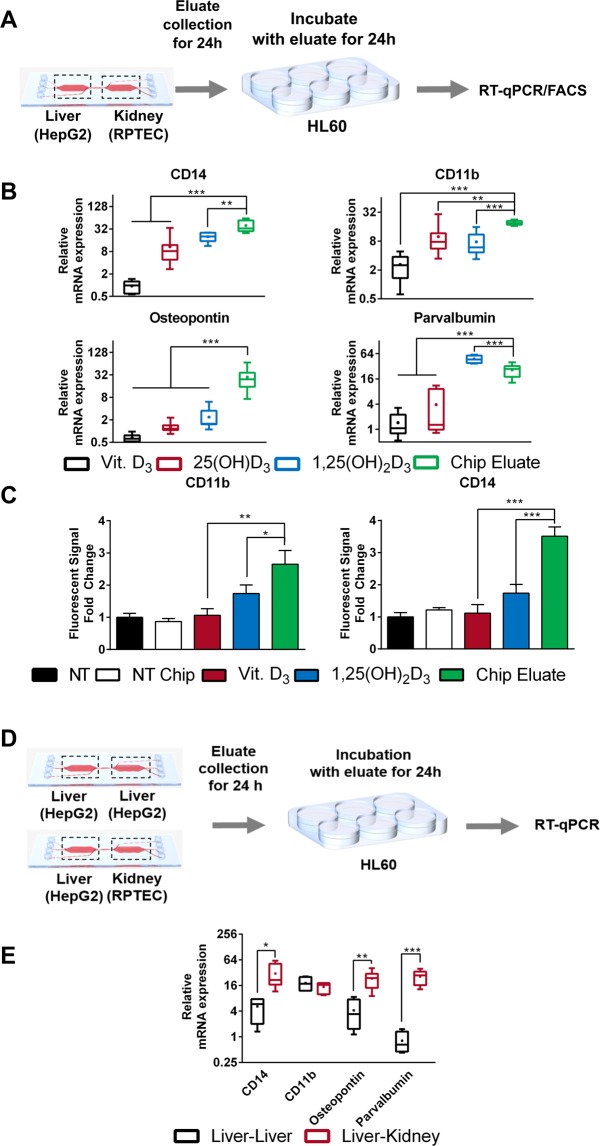
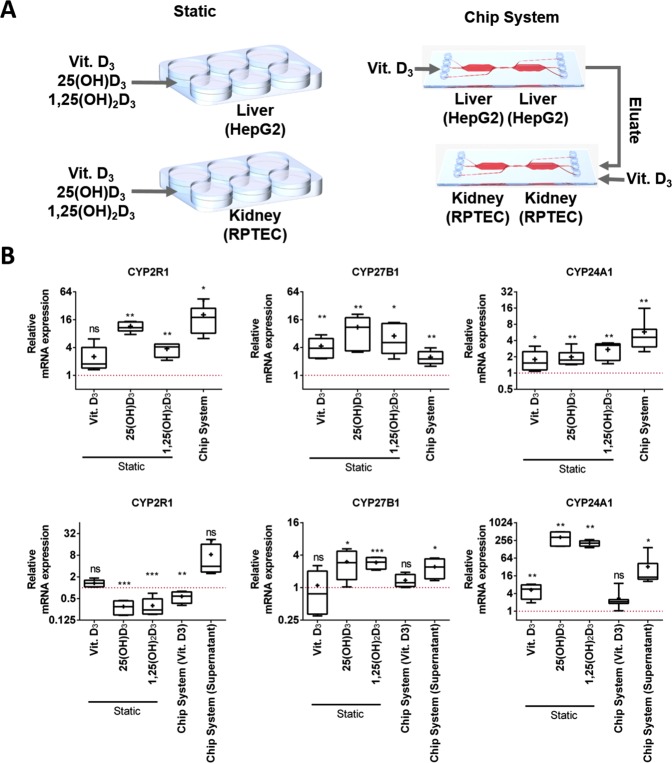
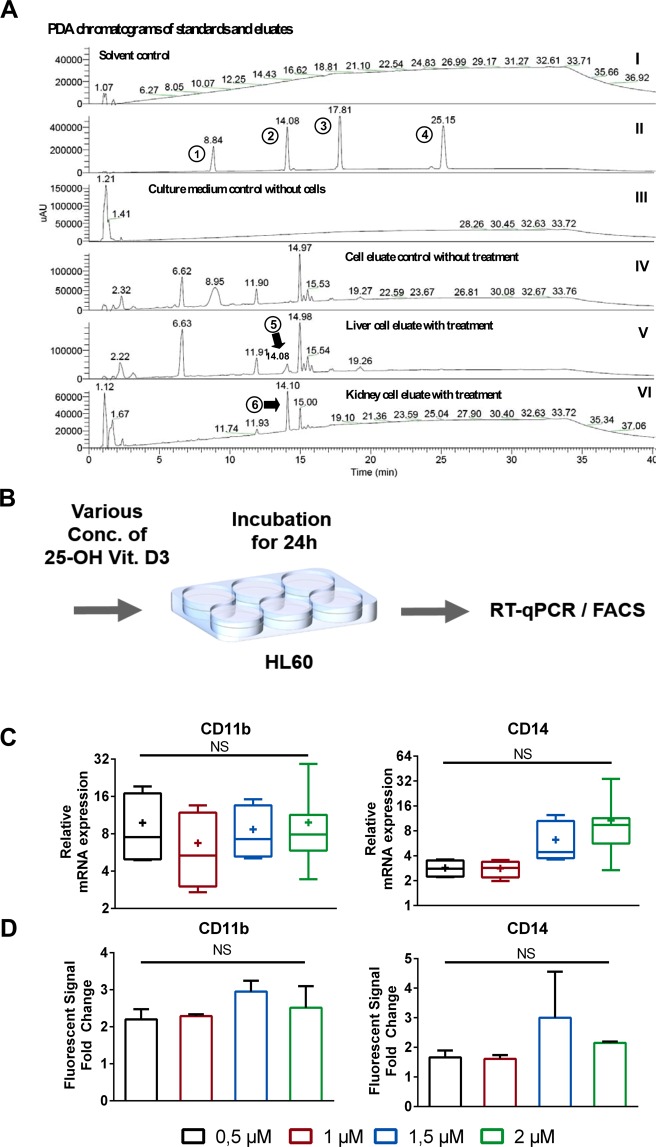
Organ-on-chip platforms provide models that allow the representation of human physiological processes in cell-based miniaturized systems. Potential pre-clinical applications include drug testing and toxicity studies. Here we describe the use of a multi-compartment micro-fluidic chip to recapitulate hepatic vitamin D metabolism (vitamin D to 25-hydroxyvitamin D) and renal bio-activation (25-hydroxyvitamin D to 1,25-dihydroxyvitamin D) in humans. In contrast to cultivation in conventional tissue culture settings, on-chip cultivation of HepG2 and RPTEC cells in interconnected chambers, used to mimic the liver and kidneys, respectively, resulted in the enhanced expression of vitamin D metabolizing enzymes (CYP2R1, CYP27B1 and CYP24A1). Pump-driven flow of vitamin D3-containing medium through the microfluidic chip produced eluate containing vitamin D3 metabolites. LC-MSMS showed a strong accumulation of 25-hydroxyvitamin D. The chip eluate induced the expression of differentiation markers in HL-60 (acute myeloid leukemia) cells, assessed by qPCR and FACS analysis, in a manner similar to treatment with reference standards indicating the presence of fully activated 1,25 dihydroxyvitamin D, although the latter was not detected in the eluate by LC-MSMS. Interestingly, 25-hydroxyvitamin D by itself led to weak activation of HL-60 cells suggesting that 25-hydroxyvitamin D is also an active metabolite. Our experiments demonstrate that complex metabolic interactions can be reconstructed outside the human body using dedicated organ-on-chip platforms. We therefore propose that such systems may be used to mimic the in vivo metabolism of various micronutrients and xenobiotics.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Abu El Maaty, M. A. et al. 1,25(OH)2D3 Disrupts glucose metabolism in prostate cancer cells leading to a truncation of the tca cycle and inhibition of txnip expression. (Biochim Biophys Acta, 2017). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
