

Longitudinal Analysis Reveals Early Development of Three MPER-Directed Neutralizing Antibody Lineages from an HIV-1-Infected Individual
- PMID: 30876875
- PMCID: PMC6555550
- DOI: 10.1016/j.immuni.2019.02.008
Longitudinal Analysis Reveals Early Development of Three MPER-Directed Neutralizing Antibody Lineages from an HIV-1-Infected Individual
Abstract
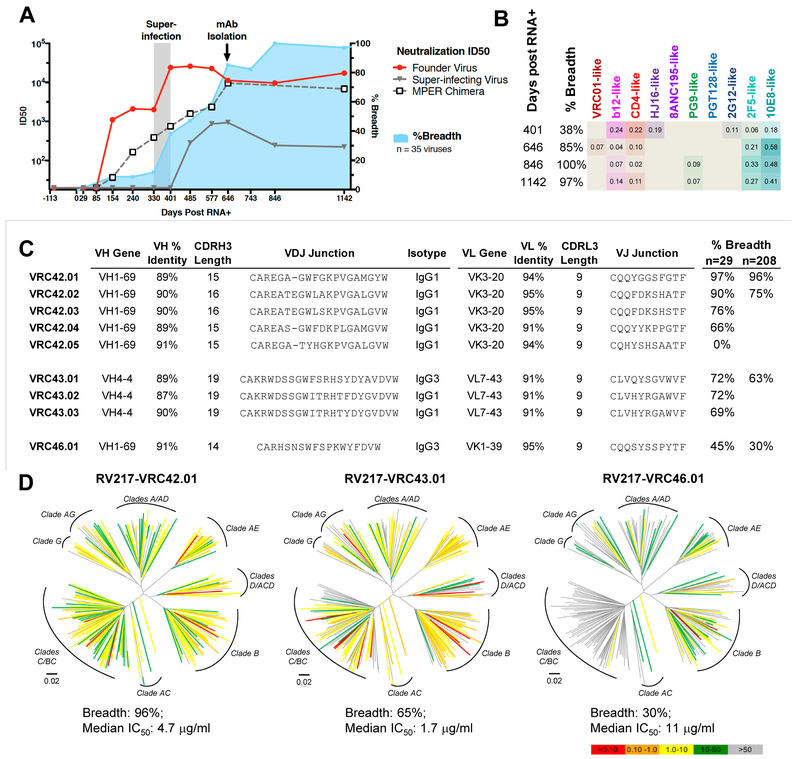
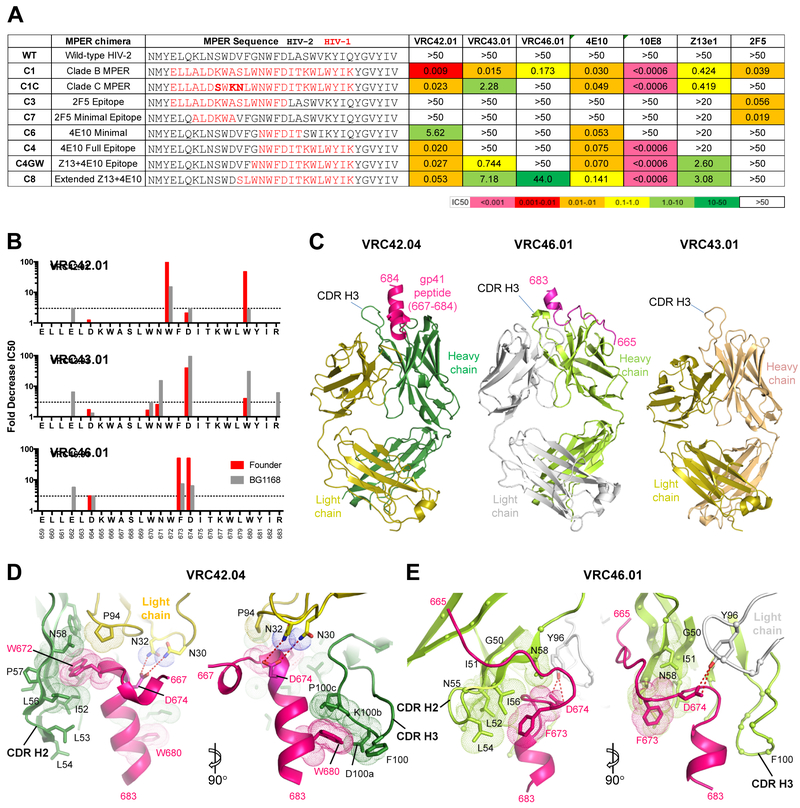
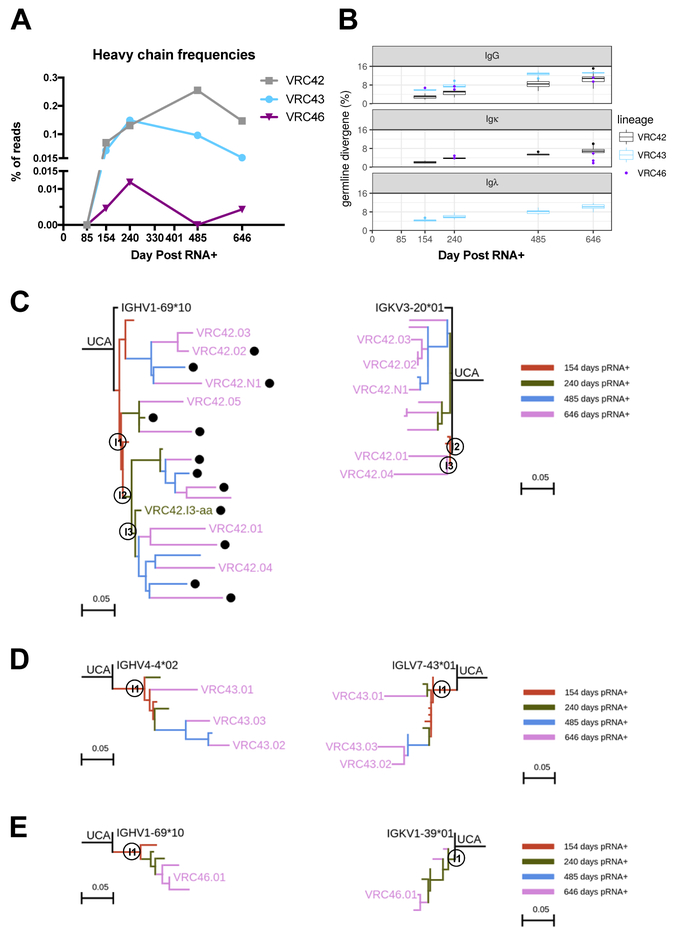
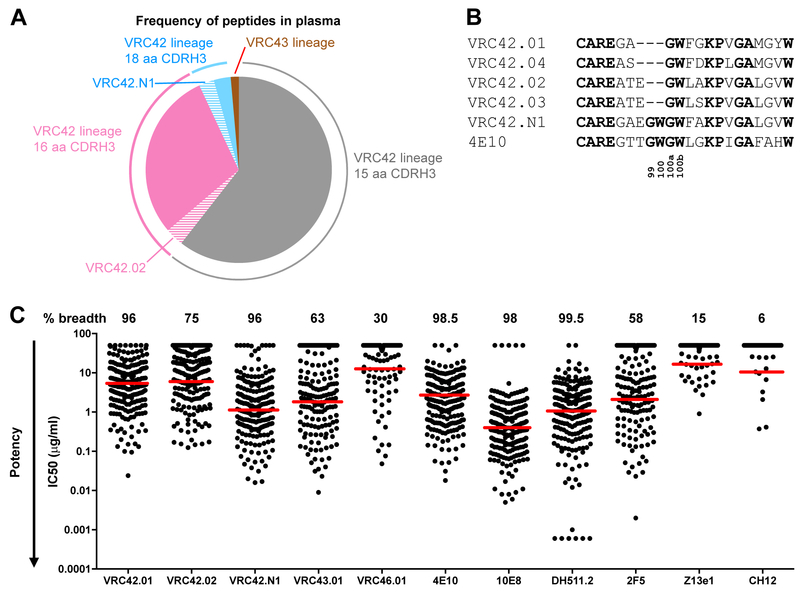
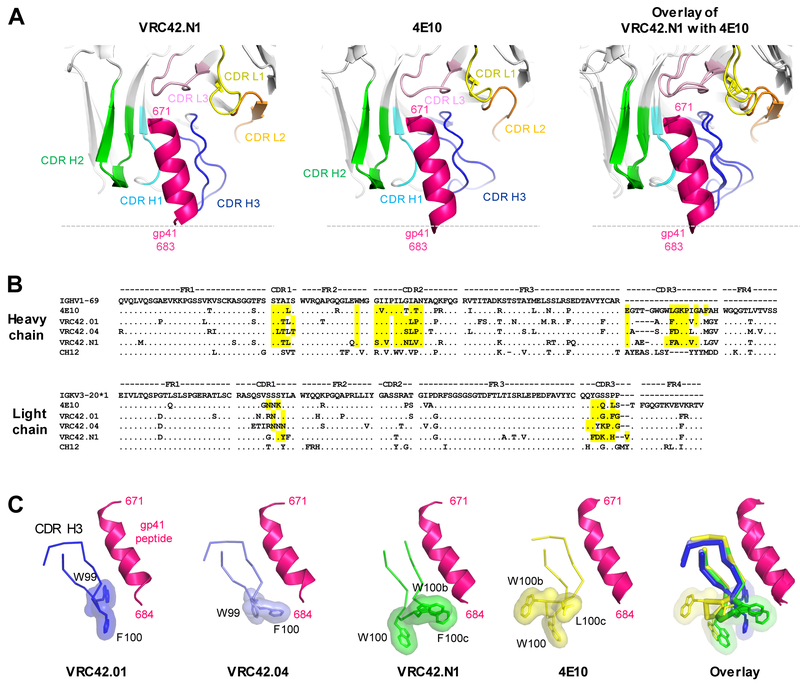
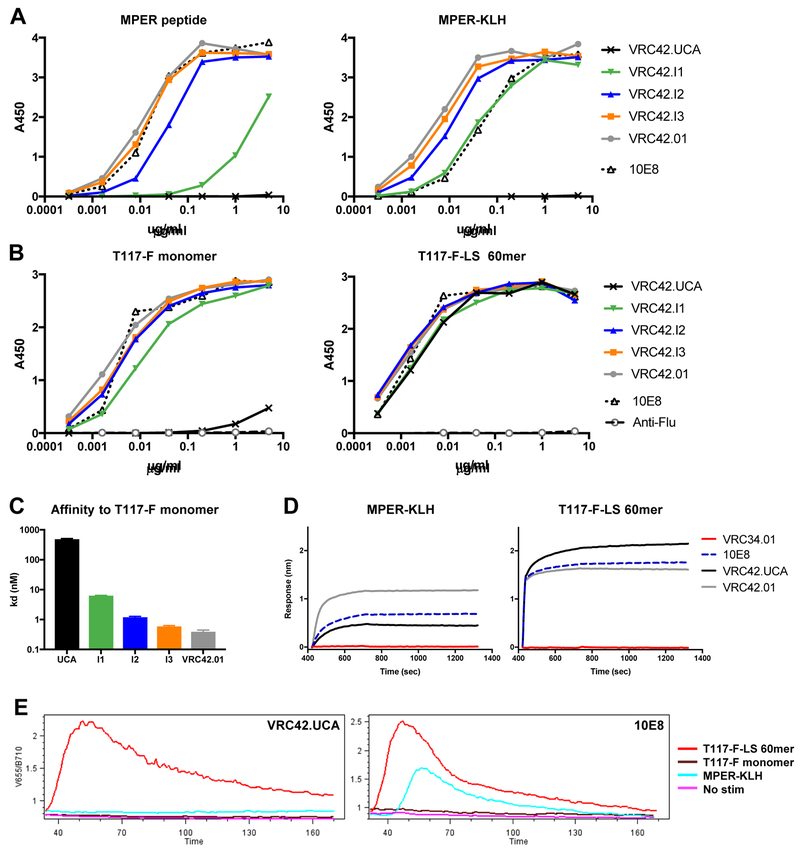
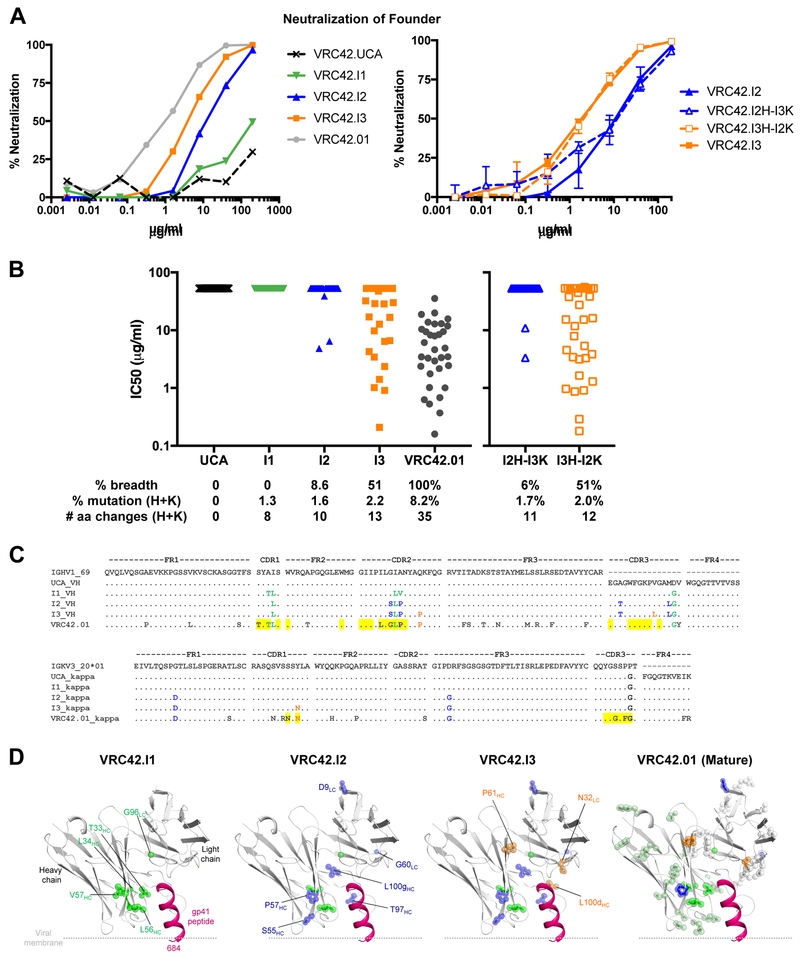
Lineage-based vaccine design is an attractive approach for eliciting broadly neutralizing antibodies (bNAbs) against HIV-1. However, most bNAb lineages studied to date have features indicative of unusual recombination and/or development. From an individual in the prospective RV217 cohort, we identified three lineages of bNAbs targeting the membrane-proximal external region (MPER) of the HIV-1 envelope. Antibodies RV217-VRC42.01, -VRC43.01, and -VRC46.01 used distinct modes of recognition and neutralized 96%, 62%, and 30%, respectively, of a 208-strain virus panel. All three lineages had modest levels of somatic hypermutation and normal antibody-loop lengths and were initiated by the founder virus MPER. The broadest lineage, VRC42, was similar to the known bNAb 4E10. A multimeric immunogen based on the founder MPER activated B cells bearing the unmutated common ancestor of VRC42, with modest maturation of early VRC42 intermediates imparting neutralization breadth. These features suggest that VRC42 may be a promising template for lineage-based vaccine design.
Keywords: HIV envelope; neutralizing antibody; next-generation sequencing.
Published by Elsevier Inc.
Figures
References
-
- Abbott RK, Lee JH, Menis S, Skog P, Rossi M, Ota T, Kulp DW, Bhullar D, Kalyuzhniy O, Havenar-Daughton C, et al. (2018). Precursor Frequency and Affinity Determine B Cell Competitive Fitness in Germinal Centers, Tested with Germline-Targeting HIV Vaccine Immunogens. Immunity 48, 133–146 e136. - PMC - PubMed
-
- Adams PD, Grosse-Kunstleve RW, Hung LW, Ioerger TR, McCoy AJ, Moriarty NW, Read RJ, Sacchettini JC, Sauter NK, and Terwilliger TC (2002). PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr D Biol Crystallogr 58, 1948–1954. - PubMed
-
- Alam SM, McAdams M, Boren D, Rak M, Scearce RM, Gao F, Camacho ZT, Gewirth D, Kelsoe G, Chen P, et al. (2007). The role of antibody polyspecificity and lipid reactivity in binding of broadly neutralizing anti-HIV-1 envelope human monoclonal antibodies 2F5 and 4E10 to glycoprotein 41 membrane proximal envelope epitopes. J Immunol 178, 4424–4435. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
