

No Evidence that Predictions and Attention Modulate the First Feedforward Sweep of Cortical Information Processing
- PMID: 30877784
- PMCID: PMC6484894
- DOI: 10.1093/cercor/bhz038
No Evidence that Predictions and Attention Modulate the First Feedforward Sweep of Cortical Information Processing
Abstract
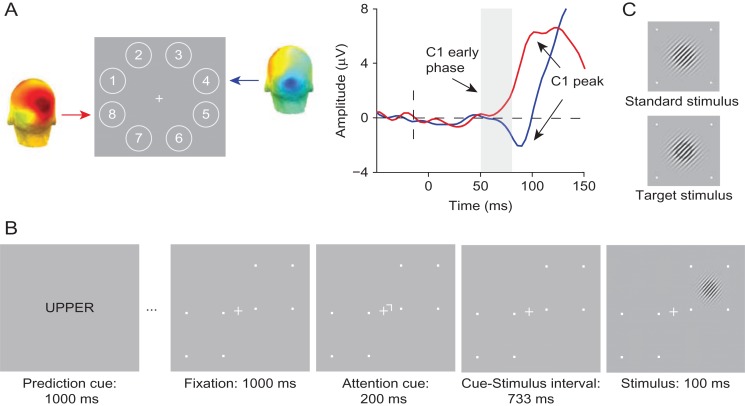
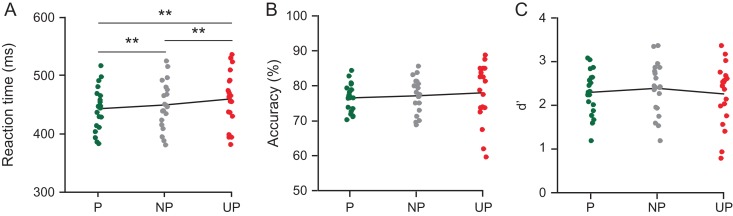
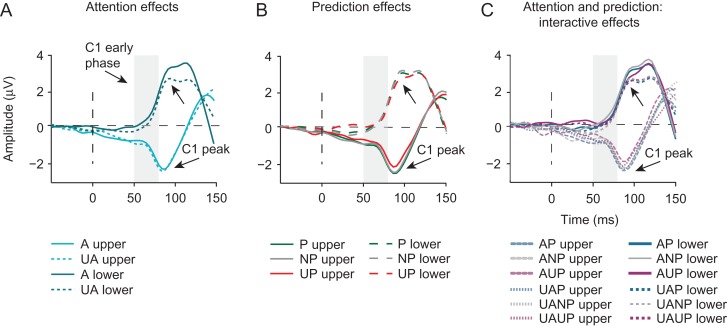
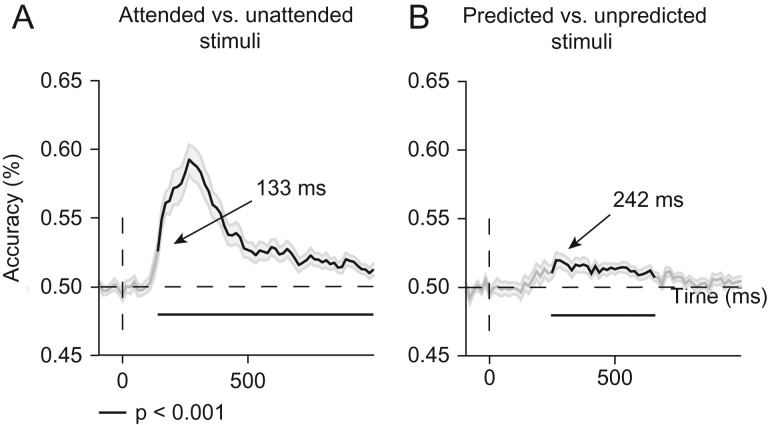
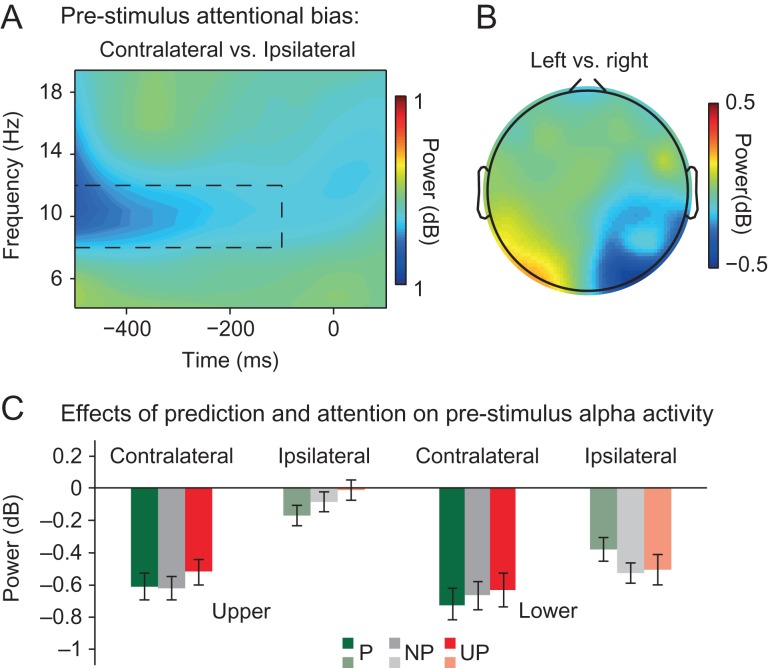
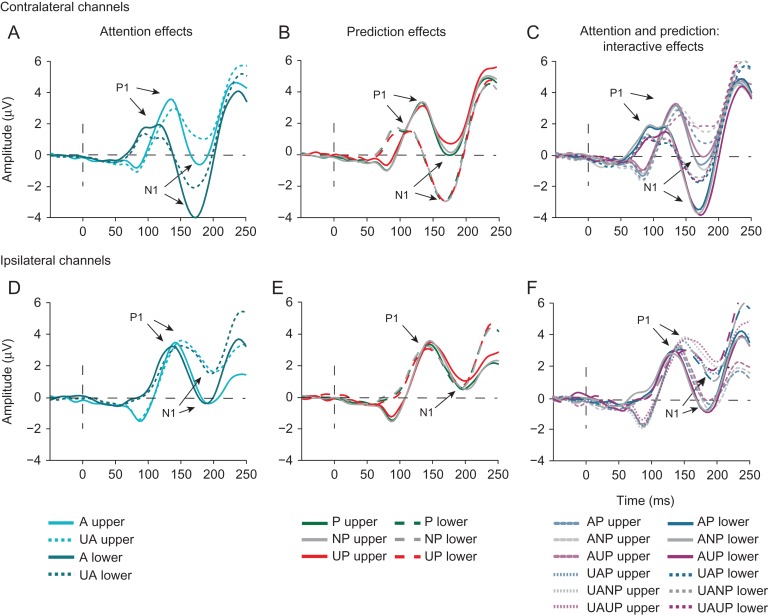
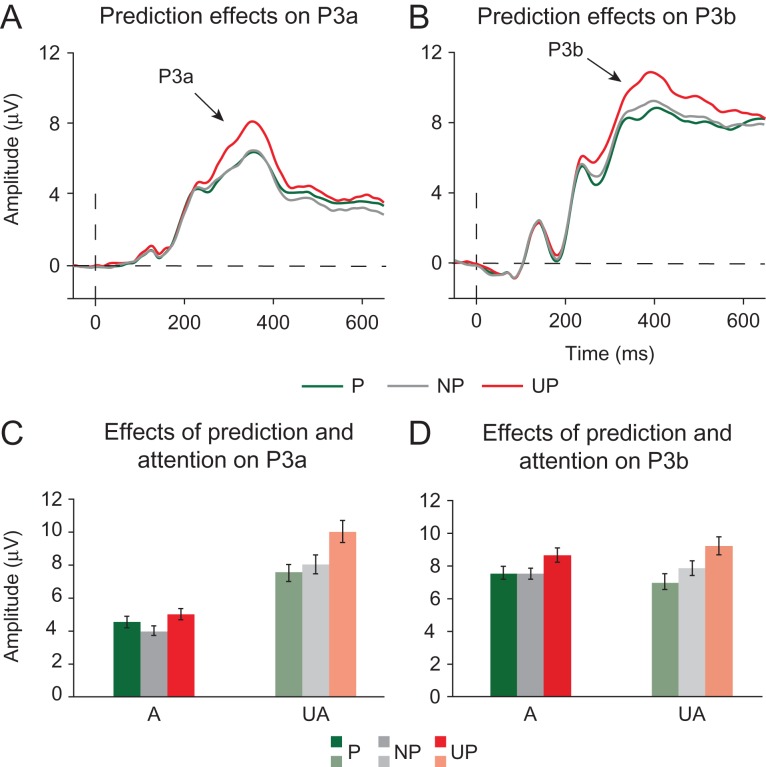
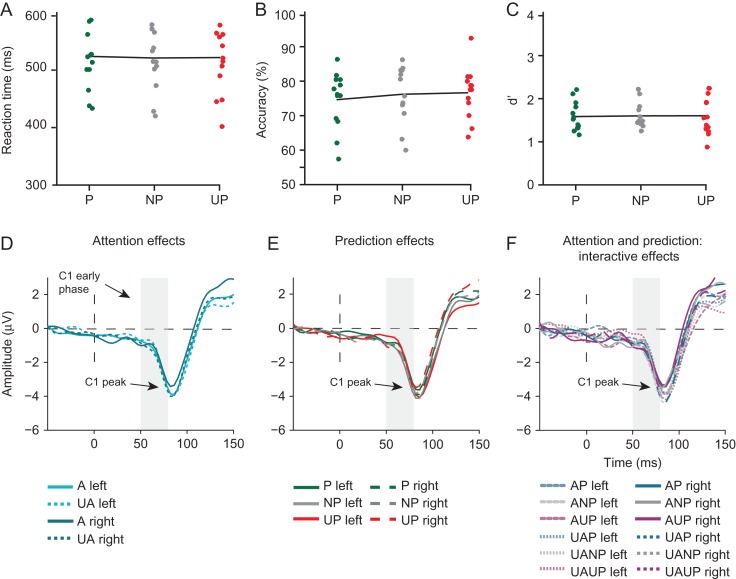
Predictive coding models propose that predictions (stimulus likelihood) reduce sensory signals as early as primary visual cortex (V1), and that attention (stimulus relevance) can modulate these effects. Indeed, both prediction and attention have been shown to modulate V1 activity, albeit with fMRI, which has low temporal resolution. This leaves it unclear whether these effects reflect a modulation of the first feedforward sweep of visual information processing and/or later, feedback-related activity. In two experiments, we used electroencephalography and orthogonally manipulated spatial predictions and attention to address this issue. Although clear top-down biases were found, as reflected in pre-stimulus alpha-band activity, we found no evidence for top-down effects on the earliest visual cortical processing stage (<80 ms post-stimulus), as indexed by the amplitude of the C1 event-related potential component and multivariate pattern analyses. These findings indicate that initial visual afferent activity may be impenetrable to top-down influences by spatial prediction and attention.
Keywords: first feedforward; human electrophysiology; prediction; spatial attention; sweep.
© The Author(s) 2019. Published by Oxford University Press.
Figures

References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
