

Co-translational, Post-translational, and Non-catalytic Roles of N-Terminal Acetyltransferases
- PMID: 30878283
- PMCID: PMC6962057
- DOI: 10.1016/j.molcel.2019.02.007
Co-translational, Post-translational, and Non-catalytic Roles of N-Terminal Acetyltransferases
Abstract
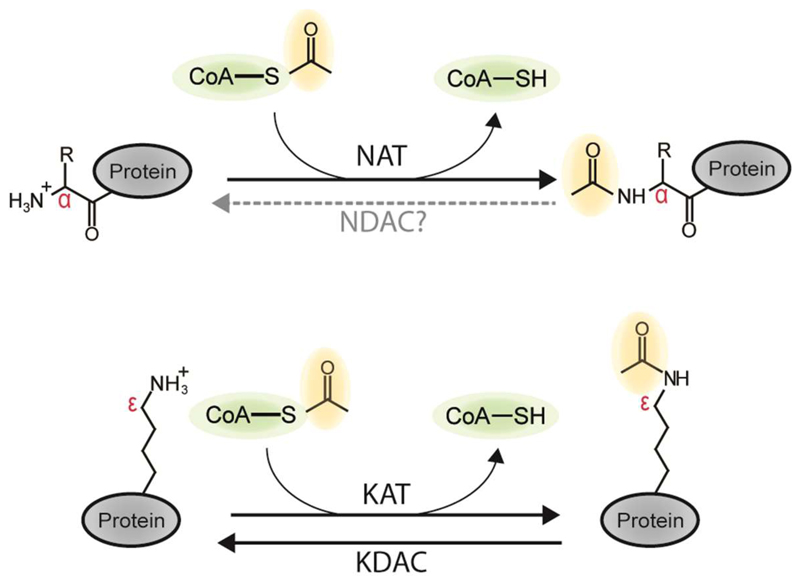
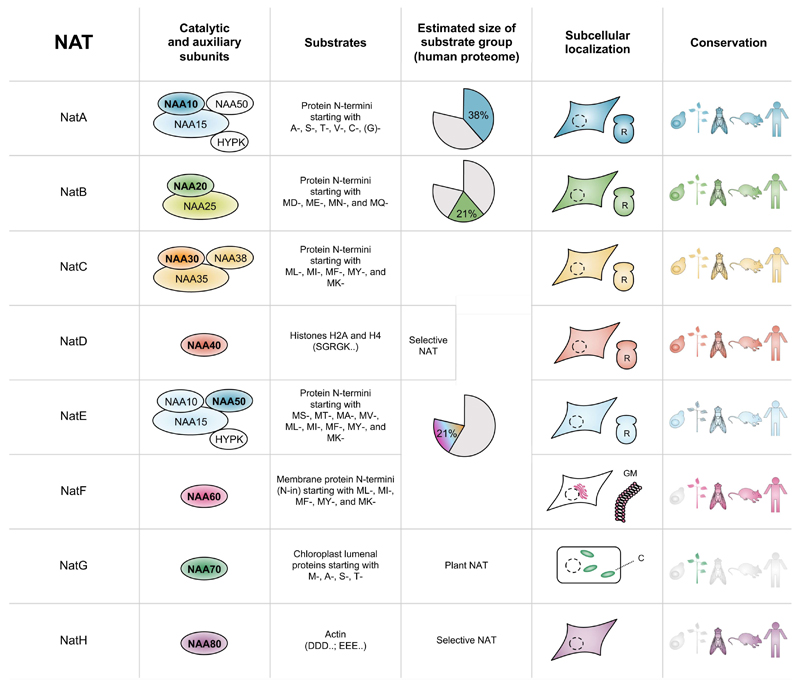
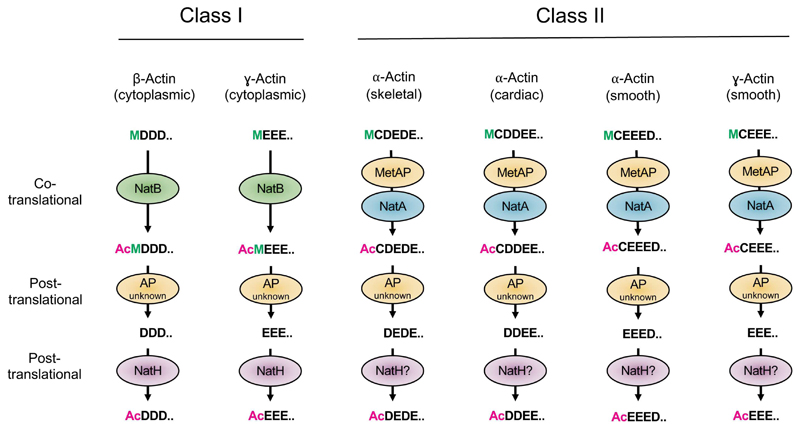
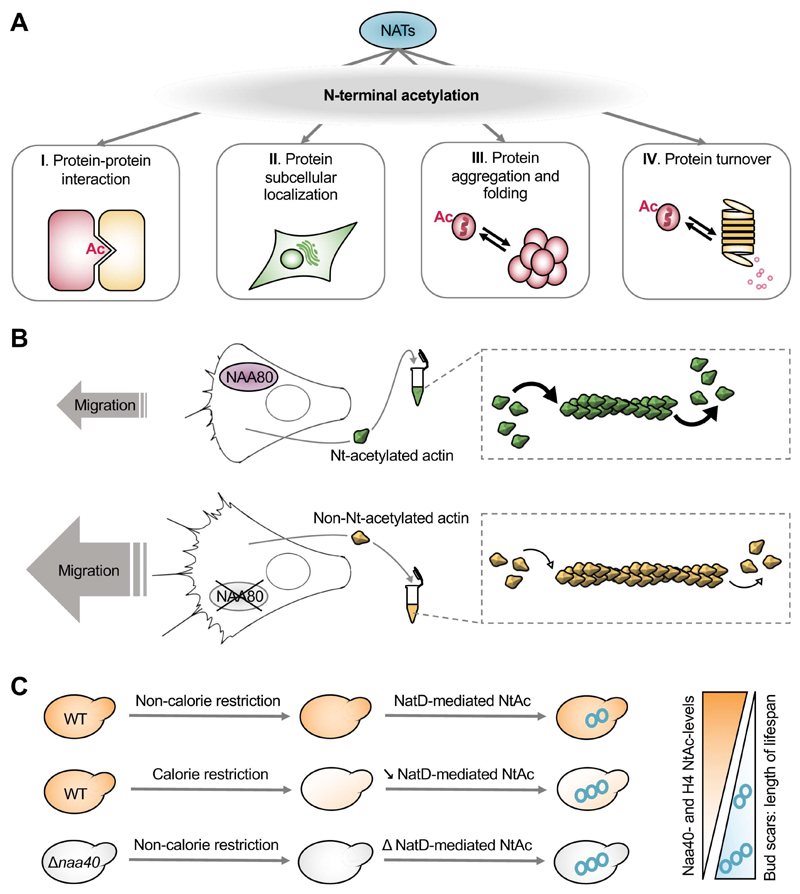
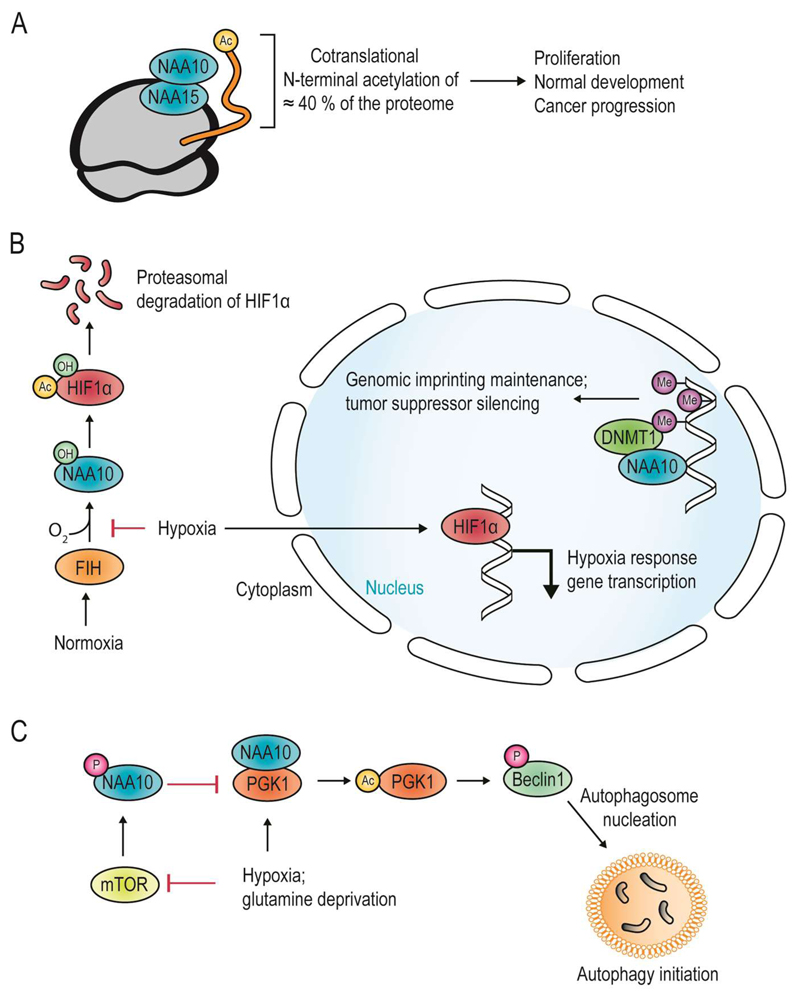
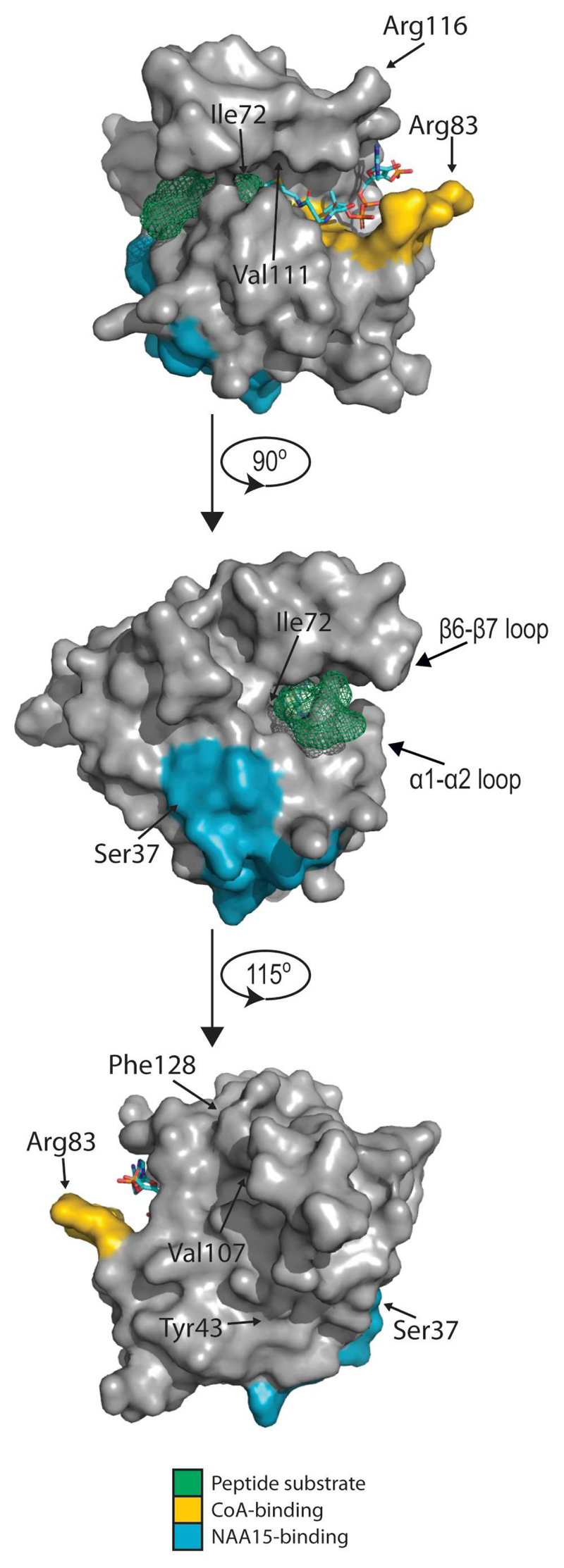
Recent studies of N-terminal acetylation have identified new N-terminal acetyltransferases (NATs) and expanded the known functions of these enzymes beyond their roles as ribosome-associated co-translational modifiers. For instance, the identification of Golgi- and chloroplast-associated NATs shows that acetylation of N termini also happens post-translationally. In addition, we now appreciate that some NATs are highly specific; for example, a dedicated NAT responsible for post-translational N-terminal acetylation of actin was recently revealed. Other studies have extended NAT function beyond Nt acetylation, including functions as lysine acetyltransferases (KATs) and non-catalytic roles. Finally, emerging studies emphasize the physiological relevance of N-terminal acetylation, including roles in calorie-restriction-induced longevity and pathological α-synuclein aggregation in Parkinson's disease. Combined, the NATs rise as multifunctional proteins, and N-terminal acetylation is gaining recognition as a major cellular regulator.
Keywords: KAT; N-terminal acetylation; N-terminal acetyltransferase; NAA10; NAA80; NAT; acetylation; actin; lysine acetyltransferase; protein modifications.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures
References
-
- Akimov V, Barrio-Hernandez I, Hansen SVF, Hallenborg P, Pedersen AK, Bekker-Jensen DB, Puglia M, Christensen SDK, Vanselow JT, Nielsen MM, et al. UbiSite approach for comprehensive mapping of lysine and N-terminal ubiquitination sites. Nature structural & molecular biology. 2018;25:631–640. - PubMed
-
- Aksnes H, Drazic A, Arnesen T. (Hyper)tension release by N-terminal acetylation. Trends in biochemical sciences. 2015a;40:422–424. - PubMed
-
- Aksnes H, Drazic A, Marie M, Arnesen T. First Things First: Vital Protein Marks by N-Terminal Acetyltransferases. Trends in biochemical sciences. 2016;41:746–760. - PubMed
-
- Aksnes H, Hole K, Arnesen T. Molecular, cellular, and physiological significance of N-terminal acetylation. International review of cell and molecular biology. 2015b;316:267–305. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
