

Single-Cell Proteomics Reveal that Quantitative Changes in Co-expressed Lineage-Specific Transcription Factors Determine Cell Fate
- PMID: 30880026
- PMCID: PMC6886472
- DOI: 10.1016/j.stem.2019.02.006
Single-Cell Proteomics Reveal that Quantitative Changes in Co-expressed Lineage-Specific Transcription Factors Determine Cell Fate
Abstract
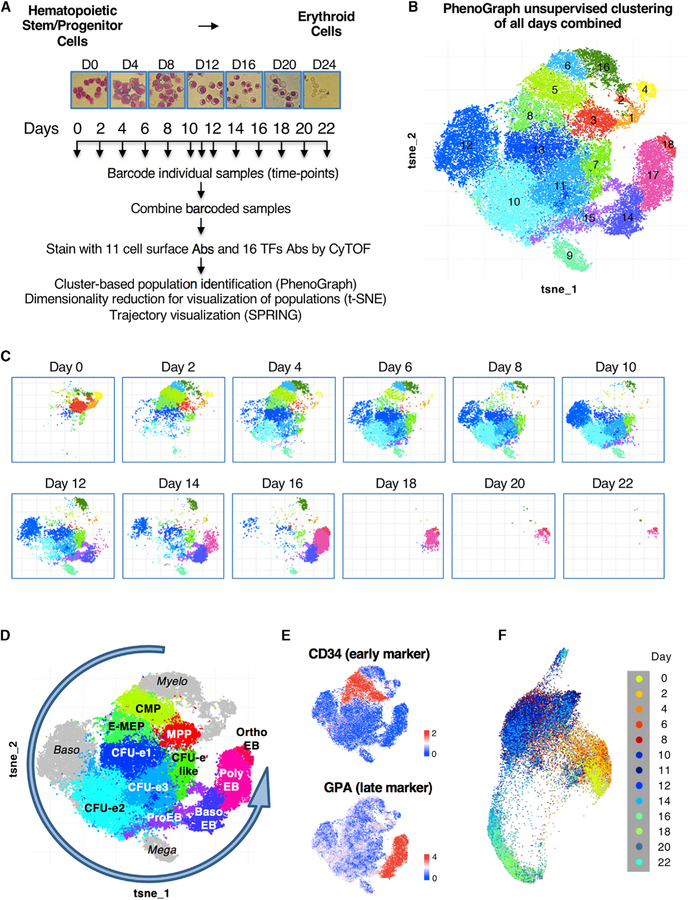
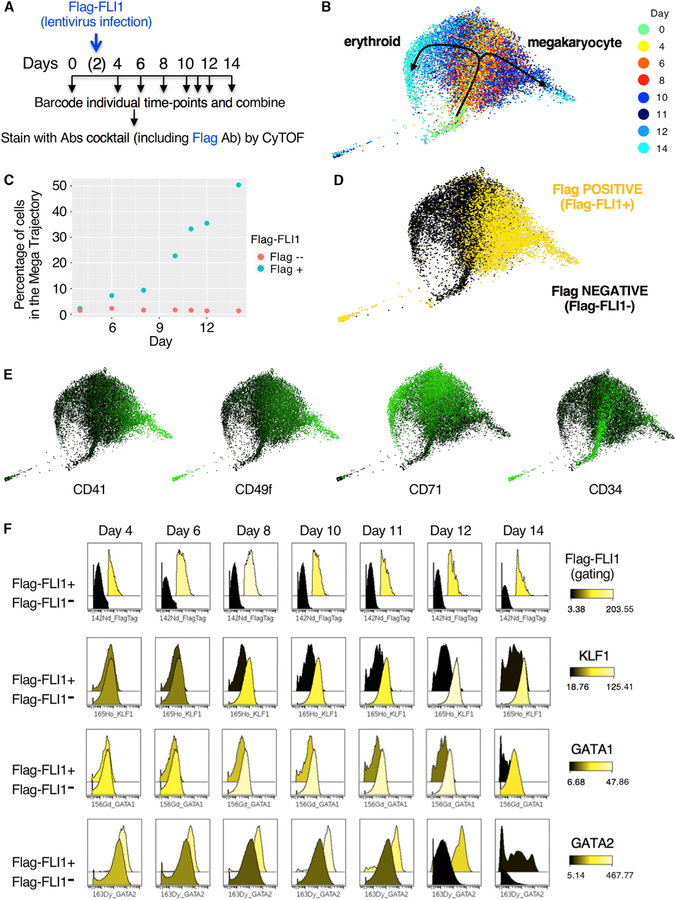
Hematopoiesis provides an accessible system for studying the principles underlying cell-fate decisions in stem cells. Proposed models of hematopoiesis suggest that quantitative changes in lineage-specific transcription factors (LS-TFs) underlie cell-fate decisions. However, evidence for such models is lacking as TF levels are typically measured via RNA expression rather than by analyzing temporal changes in protein abundance. Here, we used single-cell mass cytometry and absolute quantification by mass spectrometry to capture the temporal dynamics of TF protein expression in individual cells during human erythropoiesis. We found that LS-TFs from alternate lineages are co-expressed, as proteins, in individual early progenitor cells and quantitative changes of LS-TFs occur gradually rather than abruptly to direct cell-fate decisions. Importantly, upregulation of a megakaryocytic TF in early progenitors is sufficient to deviate cells from an erythroid to a megakaryocyte trajectory, showing that quantitative changes in protein abundance of LS-TFs in progenitors can determine alternate cell fates.
Keywords: CyTOF; FLI1; KLF1; cell fate; erythropoiesis; hematopoiesis; mass cytometry; proteomics; single cell; transcription.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures


Comment in
-
Transcription Factor Stoichiometry Drives Cell Fate: Single-Cell Proteomics to the Rescue.Cell Stem Cell. 2019 May 2;24(5):673-674. doi: 10.1016/j.stem.2019.03.002. Cell Stem Cell. 2019. PMID: 31051126
References
-
- Bouilloux F, Juban G, Cohet N, Buet D, Guyot B, Vainchenker W, Louache F, and Morlé F (2008). EKLF restricts megakaryocytic differentiation at the benefit of erythrocytic differentiation. Blood 112, 576–584. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
