

Overexpression of Dlx2 enhances osteogenic differentiation of BMSCs and MC3T3-E1 cells via direct upregulation of Osteocalcin and Alp
- PMID: 30880332
- PMCID: PMC6421343
- DOI: 10.1038/s41368-019-0046-1
Overexpression of Dlx2 enhances osteogenic differentiation of BMSCs and MC3T3-E1 cells via direct upregulation of Osteocalcin and Alp
Abstract
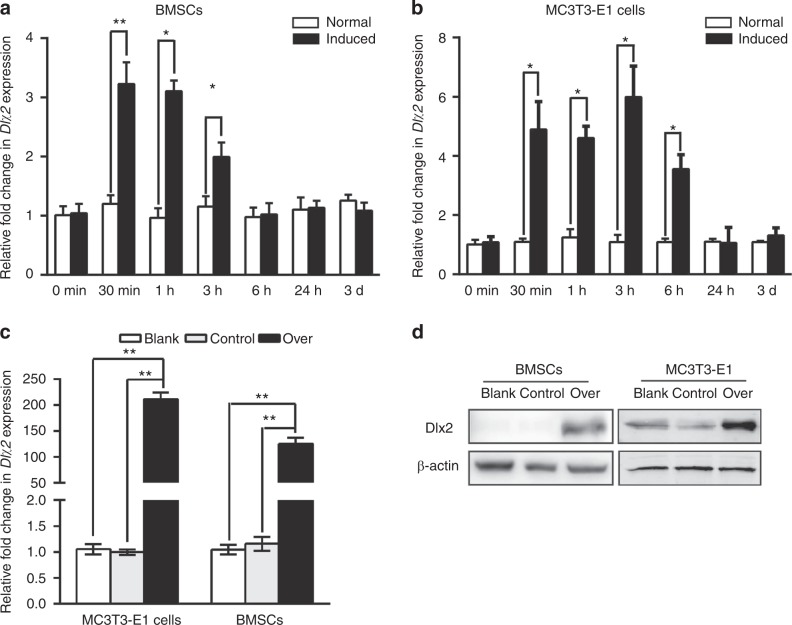
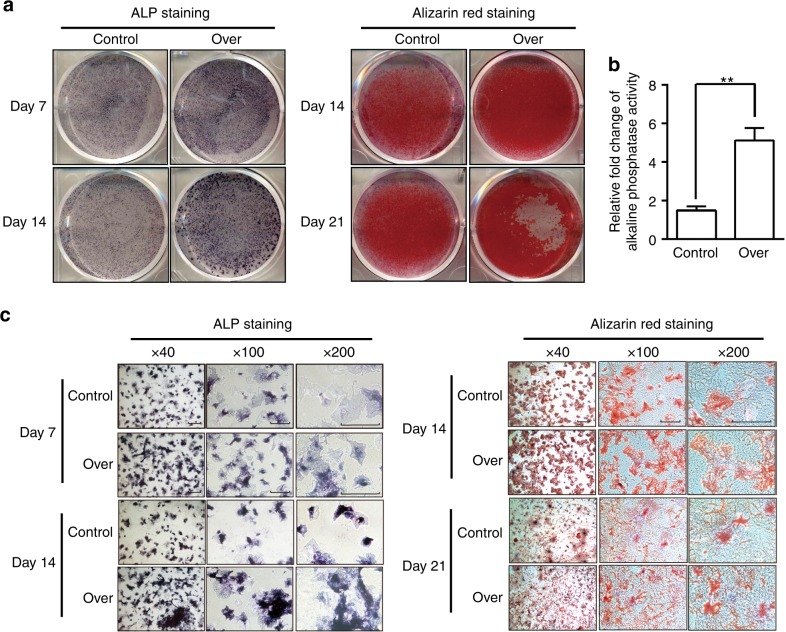
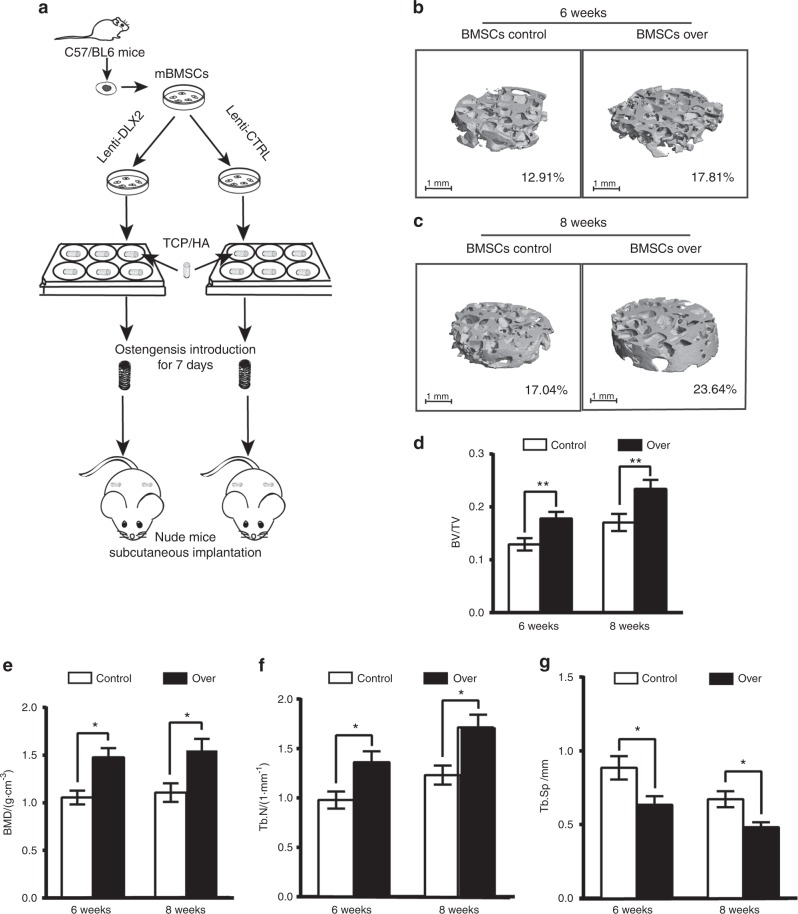
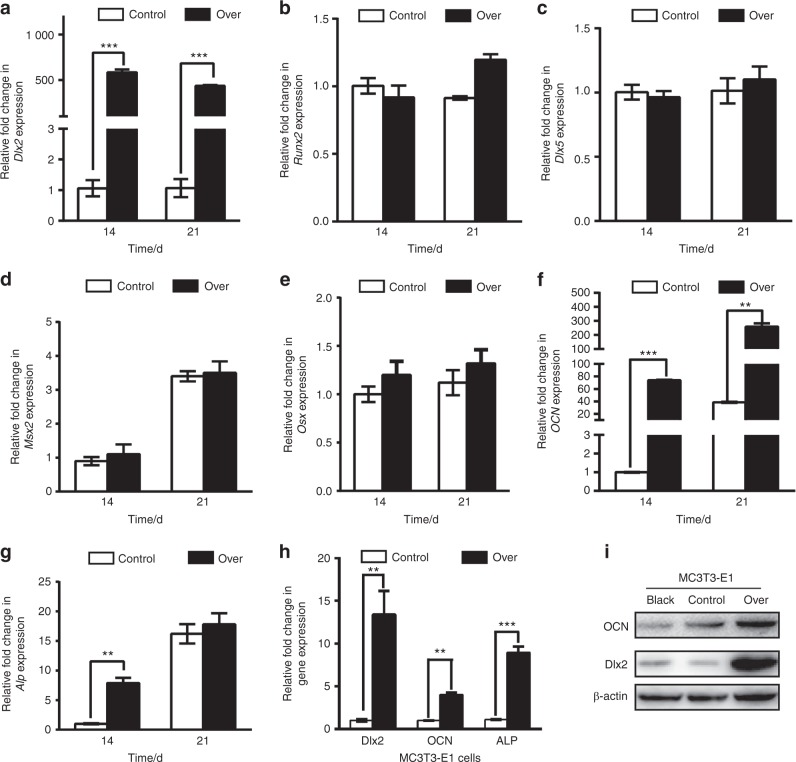
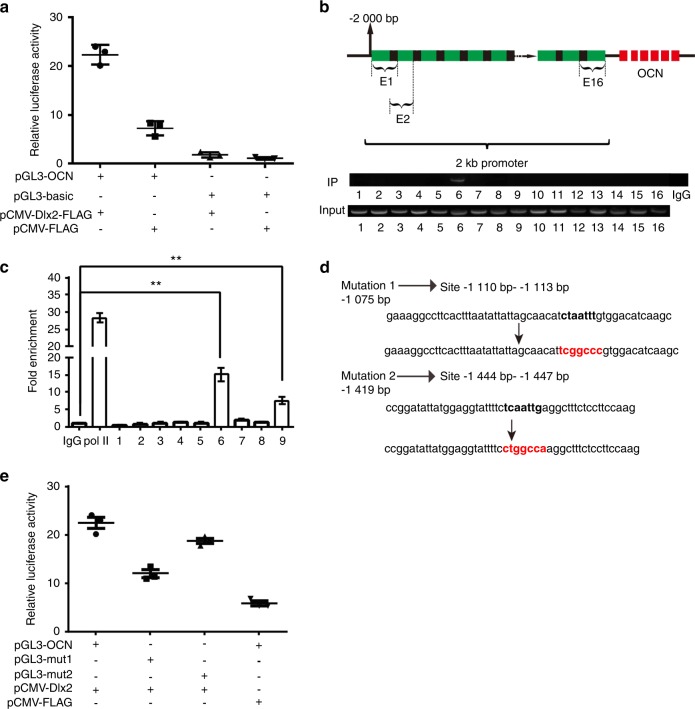
Genetic studies have revealed a critical role of Distal-homeobox (Dlx) genes in bone formation, and our previous study showed that Dlx2 overexpressing in neural crest cells leads to profound abnormalities of the craniofacial tissues. The aim of this study was to investigate the role and the underlying molecular mechanisms of Dlx2 in osteogenic differentiation of mouse bone marrow stromal cells (BMSCs) and pre-osteoblast MC3T3-E1 cells. Initially, we observed upregulation of Dlx2 during the early osteogenesis in BMSCs and MC3T3-E1 cells. Moreover, Dlx2 overexpression enhanced alkaline phosphatase (ALP) activity and extracellular matrix mineralization in BMSCs and MC3T3-E1 cell line. In addition, micro-CT of implanted tissues in nude mice confirmed that Dlx2 overexpression in BMSCs promoted bone formation in vivo. Unexpectedly, Dlx2 overexpression had little impact on the expression level of the pivotal osteogenic transcription factors Runx2, Dlx5, Msx2, and Osterix, but led to upregulation of Alp and Osteocalcin (OCN), both of which play critical roles in promoting osteoblast maturation. Importantly, luciferase analysis showed that Dlx2 overexpression stimulated both OCN and Alp promoter activity. Through chromatin-immunoprecipitation assay and site-directed mutagenesis analysis, we provide molecular evidence that Dlx2 transactivates OCN and Alp expression by directly binding to the Dlx2-response cis-acting elements in the promoter of the two genes. Based on these findings, we demonstrate that Dlx2 overexpression enhances osteogenic differentiation in vitro and accelerates bone formation in vivo via direct upregulation of the OCN and Alp gene, suggesting that Dlx2 plays a crucial role in osteogenic differentiation and bone formation.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Distal-less homeobox 2 promotes the osteogenic differentiation potential of stem cells from apical papilla.Cell Tissue Res. 2014 Jul;357(1):133-43. doi: 10.1007/s00441-014-1833-9. Epub 2014 Apr 23. Cell Tissue Res. 2014. PMID: 24756434
-
Effects of DLX2 overexpression on the osteogenic differentiation of MC3T3-E1 cells.Exp Ther Med. 2015 Jun;9(6):2173-2179. doi: 10.3892/etm.2015.2378. Epub 2015 Mar 20. Exp Ther Med. 2015. PMID: 26136955 Free PMC article.
-
miR-124 negatively regulates osteogenic differentiation and in vivo bone formation of mesenchymal stem cells.J Cell Biochem. 2015 May;116(5):730-42. doi: 10.1002/jcb.25026. J Cell Biochem. 2015. PMID: 25424317
-
Multiple functions of Dlx genes.Int J Dev Biol. 2000;44(6):619-26. Int J Dev Biol. 2000. PMID: 11061425 Review.
-
What is the function of osteocalcin?J Oral Biosci. 2020 Sep;62(3):223-227. doi: 10.1016/j.job.2020.05.004. Epub 2020 Jun 12. J Oral Biosci. 2020. PMID: 32535287 Review.
Cited by
-
Arctiin elevates osteogenic differentiation of MC3T3-E1 cells by modulating cyclin D1.Bioengineered. 2022 Apr;13(4):10866-10874. doi: 10.1080/21655979.2022.2066047. Bioengineered. 2022. PMID: 35473505 Free PMC article.
-
Effects of Dlx2 overexpression on the genes associated with the maxillary process in the early mouse embryo.Front Genet. 2023 Feb 20;14:1085263. doi: 10.3389/fgene.2023.1085263. eCollection 2023. Front Genet. 2023. PMID: 36891149 Free PMC article.
-
Nanotopographical 3D-Printed Poly(ε-caprolactone) Scaffolds Enhance Proliferation and Osteogenic Differentiation of Urine-Derived Stem Cells for Bone Regeneration.Pharmaceutics. 2022 Jul 8;14(7):1437. doi: 10.3390/pharmaceutics14071437. Pharmaceutics. 2022. PMID: 35890332 Free PMC article.
-
Strategies for the Covalent Anchoring of a BMP-2-Mimetic Peptide to PEEK Surface for Bone Tissue Engineering.Materials (Basel). 2023 May 21;16(10):3869. doi: 10.3390/ma16103869. Materials (Basel). 2023. PMID: 37241496 Free PMC article.
-
CH02 peptide-stimulated periodontal ligament cells enhance periodontal defect repair in rats.BMC Oral Health. 2025 Jul 3;25(1):1078. doi: 10.1186/s12903-025-06393-5. BMC Oral Health. 2025. PMID: 40611174 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
