

A unique aspartyl protease gene expansion in Talaromyces marneffei plays a role in growth inside host phagocytes
- PMID: 30880596
- PMCID: PMC6527018
- DOI: 10.1080/21505594.2019.1593776
A unique aspartyl protease gene expansion in Talaromyces marneffei plays a role in growth inside host phagocytes
Abstract
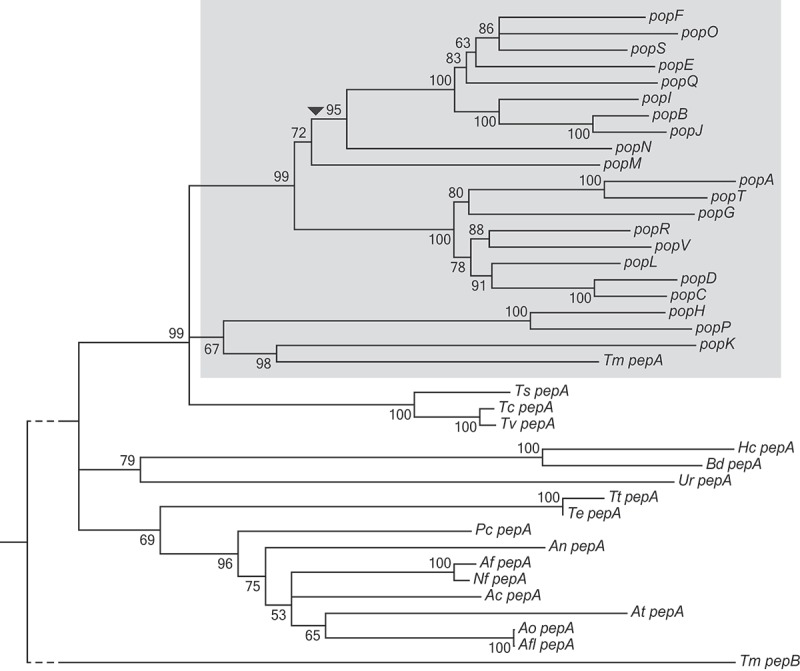
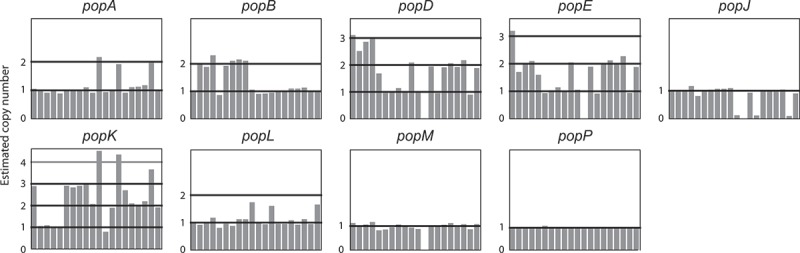
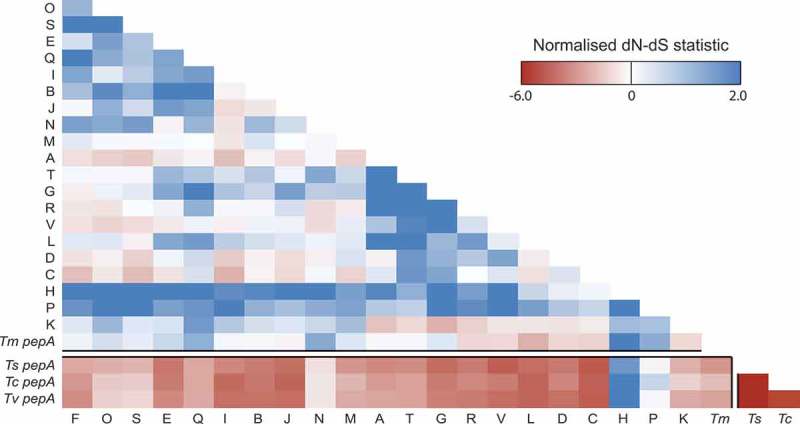
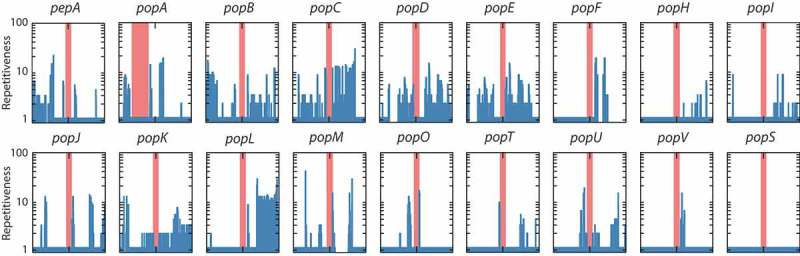
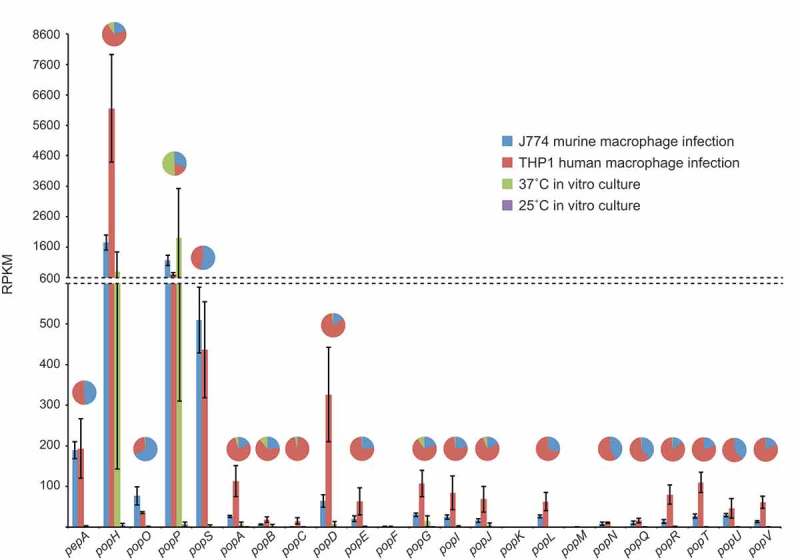
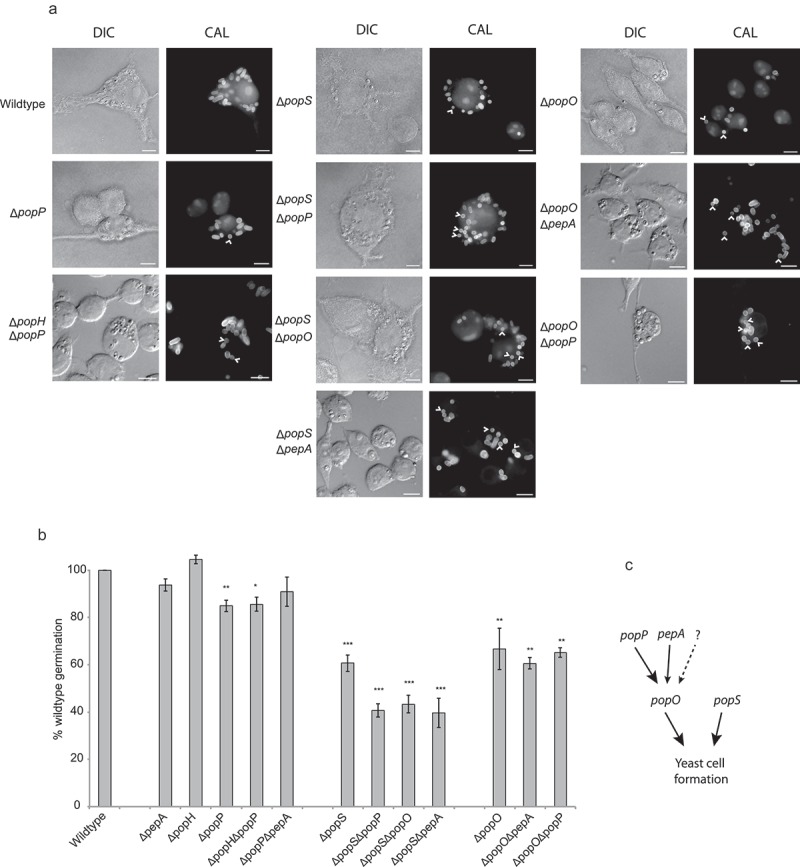
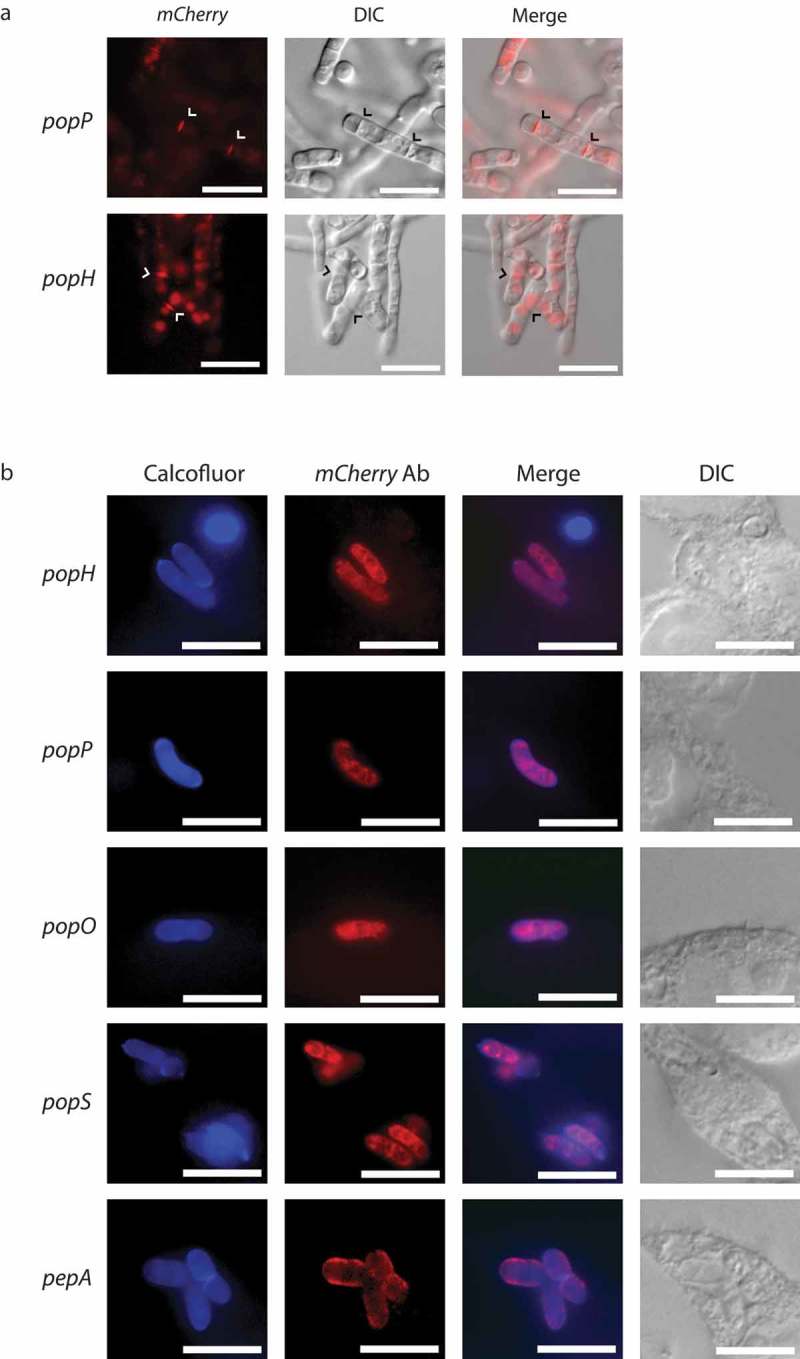
Aspartyl proteases are a widely represented class of proteolytic enzymes found in eukaryotes and retroviruses. They have been associated with pathogenicity in a range of disease-causing microorganisms. The dimorphic human-pathogenic fungus Talaromyces marneffei has a large expansion of these proteases identified through genomic analyses. Here we characterize the expansion of these genes (pop - paralogue of pep) and their role in T. marneffei using computational and molecular approaches. Many of the genes in this monophyletic family show copy number variation and positive selection despite the preservation of functional regions and possible redundancy. We show that the expression profile of these genes differs and some are expressed during intracellular growth in the host. Several of these proteins have distinctive localization as well as both additive and epistatic effects on the formation of yeast cells during macrophage infections. The data suggest that this is a recently evolved aspartyl protease gene family which affects intracellular growth and contributes to the pathogenicity of T. marneffei.
Keywords: aspartyl protease; evolution; pathogenic fungi.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous