

Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli
- PMID: 30881364
- PMCID: PMC6407669
- DOI: 10.3389/fpls.2019.00157
Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli
Erratum in
-
Corrigendum: Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli.Front Plant Sci. 2019 Apr 9;10:420. doi: 10.3389/fpls.2019.00420. eCollection 2019. Front Plant Sci. 2019. PMID: 31024593 Free PMC article.
Abstract
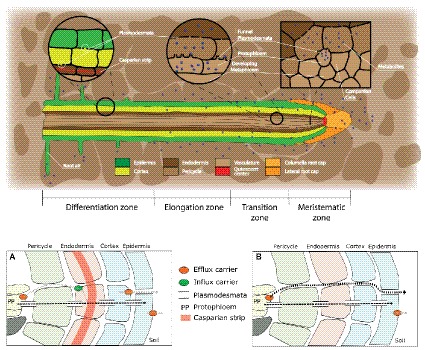
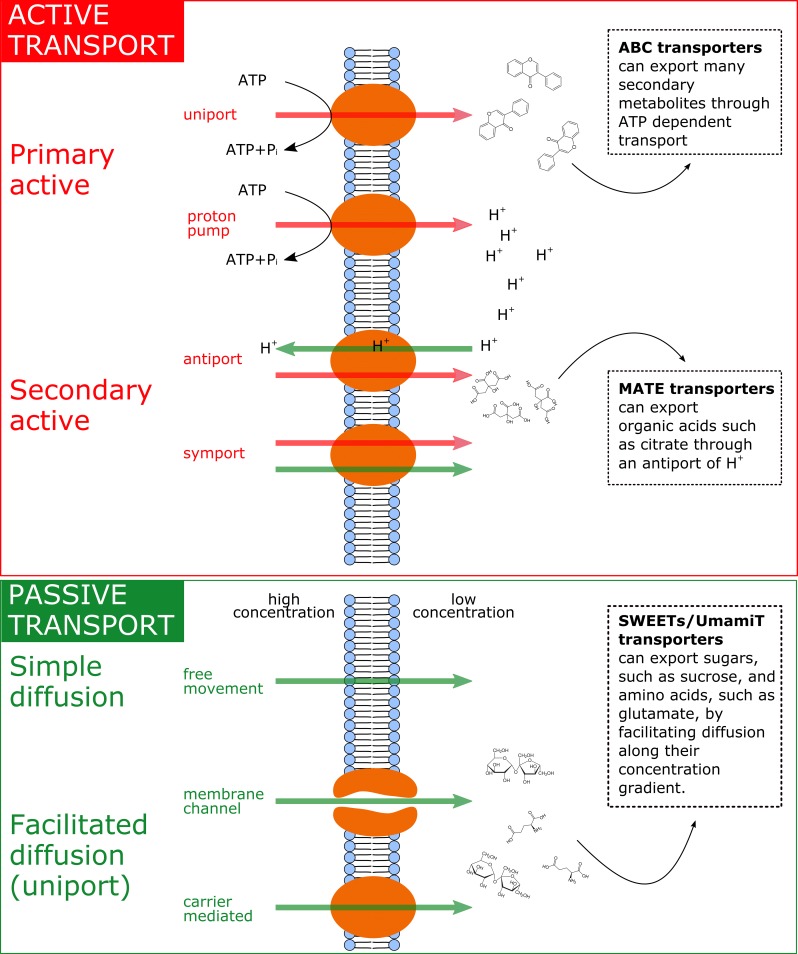
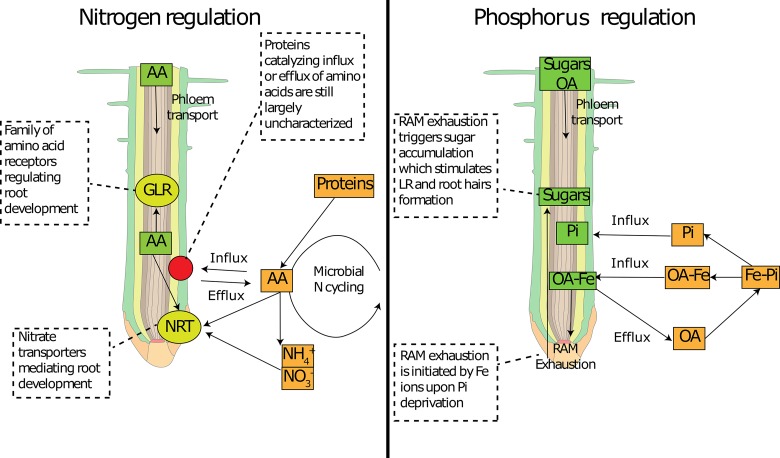
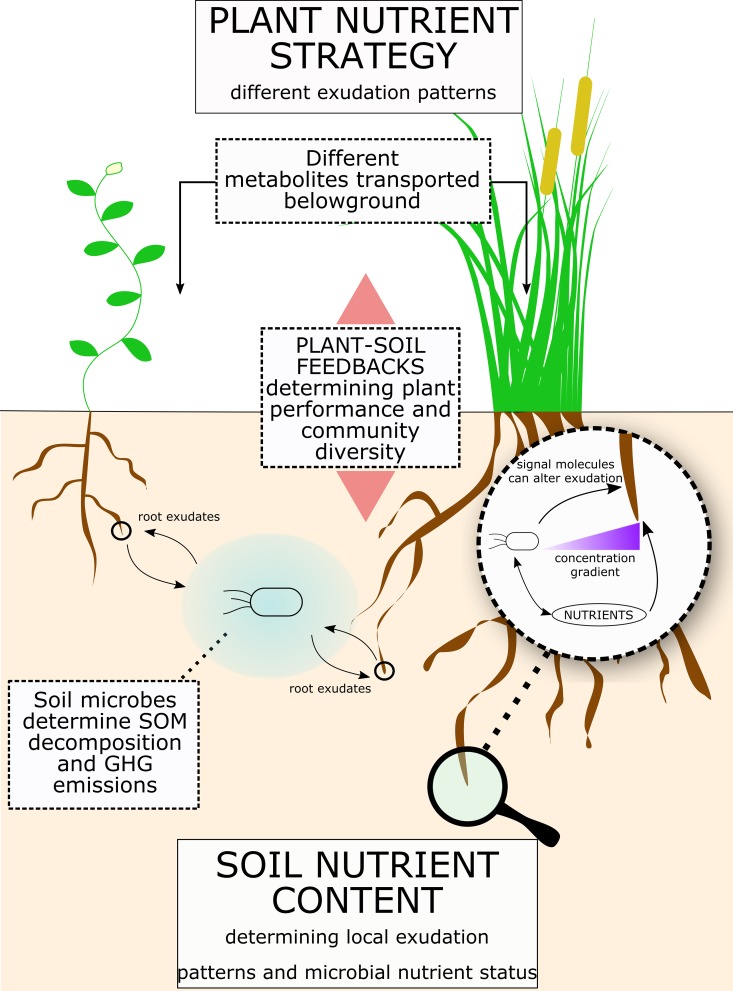
Root exudation is an important process determining plant interactions with the soil environment. Many studies have linked this process to soil nutrient mobilization. Yet, it remains unresolved how exudation is controlled and how exactly and under what circumstances plants benefit from exudation. The majority of root exudates including primary metabolites (sugars, amino acids, and organic acids) are believed to be passively lost from the root and used by rhizosphere-dwelling microbes. In this review, we synthetize recent advances in ecology and plant biology to explain and propose mechanisms by which root exudation of primary metabolites is controlled, and what role their exudation plays in plant nutrient acquisition strategies. Specifically, we propose a novel conceptual framework for root exudates. This framework is built upon two main concepts: (1) root exudation of primary metabolites is driven by diffusion, with plants and microbes both modulating concentration gradients and therefore diffusion rates to soil depending on their nutritional status; (2) exuded metabolite concentrations can be sensed at the root tip and signals are translated to modify root architecture. The flux of primary metabolites through root exudation is mostly located at the root tip, where the lack of cell differentiation favors diffusion of metabolites to the soil. We show examples of how the root tip senses concentration changes of exuded metabolites and translates that into signals to modify root growth. Plants can modify the concentration of metabolites either by controlling source/sink processes or by expressing and regulating efflux carriers, therefore challenging the idea of root exudation as a purely unregulated passive process. Through root exudate flux, plants can locally enhance concentrations of many common metabolites, which can serve as sensors and integrators of the plant nutritional status and of the nutrient availability in the surrounding environment. Plant-associated micro-organisms also constitute a strong sink for plant carbon, thereby increasing concentration gradients of metabolites and affecting root exudation. Understanding the mechanisms of and the effects that environmental stimuli have on the magnitude and type of root exudation will ultimately improve our knowledge of processes determining soil CO2 emissions, ecosystem functioning, and how to improve the sustainability of agricultural production.
Keywords: mycorrhiza; nutrient sensing; priming effect; root architecture; root exudates; soil micro-organisms.
Figures
Similar articles
-
The Sorghum bicolor Root Exudate Sorgoleone Shapes Bacterial Communities and Delays Network Formation.mSystems. 2021 Mar 16;6(2):e00749-20. doi: 10.1128/mSystems.00749-20. mSystems. 2021. PMID: 33727394 Free PMC article.
-
Cover crop root exudates impact soil microbiome functional trajectories in agricultural soils.Microbiome. 2024 Sep 28;12(1):183. doi: 10.1186/s40168-024-01886-x. Microbiome. 2024. PMID: 39342284 Free PMC article.
-
A dynamic rhizosphere interplay between tree roots and soil bacteria under drought stress.Elife. 2022 Jul 20;11:e79679. doi: 10.7554/eLife.79679. Elife. 2022. PMID: 35858113 Free PMC article.
-
Metabolic niches in the rhizosphere microbiome: dependence on soil horizons, root traits and climate variables in forest ecosystems.Front Plant Sci. 2024 Apr 5;15:1344205. doi: 10.3389/fpls.2024.1344205. eCollection 2024. Front Plant Sci. 2024. PMID: 38645395 Free PMC article. Review.
-
Plant root exudation under drought: implications for ecosystem functioning.New Phytol. 2020 Mar;225(5):1899-1905. doi: 10.1111/nph.16223. Epub 2019 Oct 26. New Phytol. 2020. PMID: 31571220 Review.
Cited by
-
Bioaugmentation of Entomopathogenic Fungi for Sustainable Agriotes Larvae (Wireworms) Management in Maize.Front Plant Sci. 2020 Sep 17;11:535005. doi: 10.3389/fpls.2020.535005. eCollection 2020. Front Plant Sci. 2020. PMID: 33042172 Free PMC article.
-
Nitrogen Uptake in Plants: The Plasma Membrane Root Transport Systems from a Physiological and Proteomic Perspective.Plants (Basel). 2021 Apr 1;10(4):681. doi: 10.3390/plants10040681. Plants (Basel). 2021. PMID: 33916130 Free PMC article. Review.
-
Nutrient and Microbiome-Mediated Plant-Soil Feedback in Domesticated and Wild Andropogoneae: Implications for Agroecosystems.Microorganisms. 2023 Dec 13;11(12):2978. doi: 10.3390/microorganisms11122978. Microorganisms. 2023. PMID: 38138123 Free PMC article.
-
A Medicago truncatula Metabolite Atlas Enables the Visualization of Differential Accumulation of Metabolites in Root Tissues.Metabolites. 2021 Apr 13;11(4):238. doi: 10.3390/metabo11040238. Metabolites. 2021. PMID: 33924579 Free PMC article.
-
Metabolic Alterations in Pisum sativum Roots during Plant Growth and Arbuscular Mycorrhiza Development.Plants (Basel). 2021 May 21;10(6):1033. doi: 10.3390/plants10061033. Plants (Basel). 2021. PMID: 34063836 Free PMC article.
References
-
- Alegria Terrazas R., Giles C., Paterson E., Robertson-Albertyn S., Cesco S., Mimmo T., et al. (2016). “Chapter one: Plant–microbiota interactions as a driver of the mineral turnover in the rhizosphere,” in Advances in applied microbiology. eds. Sariaslani S., Gadd G. M. (Cambridge, Massachusetts, USA: Academic Press; ), 1–67. - PubMed
-
- Badri D. V., Loyola-Vargas V. M., Du J., Stermitz F. R., Broeckling C. D., Iglesias-Andreu L., et al. . (2008). Transcriptome analysis of Arabidopsis roots treated with signaling compounds: a focus on signal transduction, metabolic regulation and secretion. New Phytol. 179, 209–223. 10.1111/j.1469-8137.2008.02458.x, PMID: - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources