

The regulatory network of Vibrio parahaemolyticus type VI secretion system 1
- PMID: 30882997
- PMCID: PMC6618117
- DOI: 10.1111/1462-2920.14594
The regulatory network of Vibrio parahaemolyticus type VI secretion system 1
Abstract
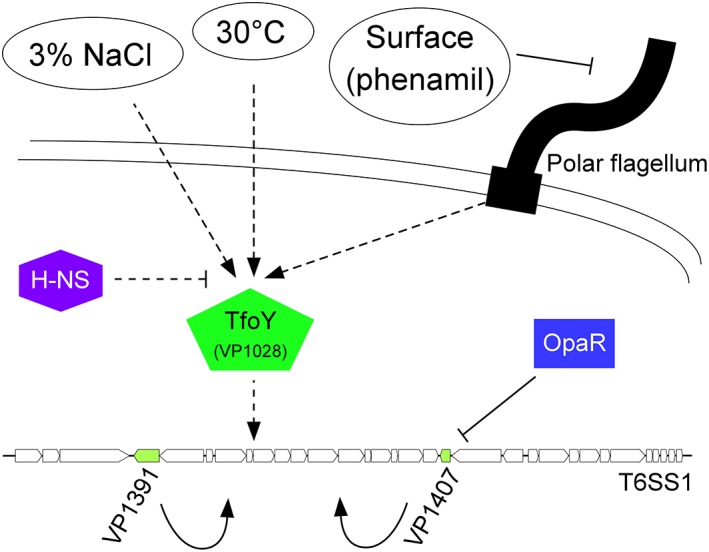
Type VI secretion systems (T6SSs) are widespread, tightly regulated, protein delivery apparatuses used by Gram-negative bacteria to outcompete their neighbours. The pathogen, Vibrio parahaemolyticus, encodes two T6SSs. These T6SSs are differentially regulated by external conditions. T6SS1, an antibacterial system predominantly found in pathogenic isolates, requires warm marine-like conditions and surface sensing for activation. The regulatory network that governs this activation is not well understood. In this work, we devised a screening methodology that allows us to easily monitor the outcome of bacterial competitions and thus to identify mutants that are defective in T6SS1-mediated bacterial killing. The methodology, termed Bacterial Competition Fluorescence (BaCoF), relies on detection of a fluorescent signal as an indicator of the survival and growth of a T6SS-sensitive, GFP-expressing prey that has been co-cultured with mutants derived from a T6SS+ attacker of interest. Using BaCoF, we screened a random transposon insertion mutant library and identified genes required for V. parahaemolyticus T6SS1 activation, among them TfoY and Tmk. We used epistasis experiments to determine the relationships between the newly identified components and other regulators that were previously described. Thus, we present here a detailed biological understanding of the T6SS1 regulatory network.
© 2019 The Authors. Environmental Microbiology published by Society for Applied Microbiology and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures





Comment in
-
Should I kill or should I go: T6SS regulation networks in Vibrio.Environ Microbiol. 2020 Jan;22(1):1-4. doi: 10.1111/1462-2920.14830. Epub 2019 Oct 25. Environ Microbiol. 2020. PMID: 31621177 No abstract available.
Similar articles
-
Vibrio parahaemolyticus type VI secretion system 1 is activated in marine conditions to target bacteria, and is differentially regulated from system 2.PLoS One. 2013 Apr 16;8(4):e61086. doi: 10.1371/journal.pone.0061086. Print 2013. PLoS One. 2013. PMID: 23613791 Free PMC article.
-
Multiple T6SSs, Mobile Auxiliary Modules, and Effectors Revealed in a Systematic Analysis of the Vibrio parahaemolyticus Pan-Genome.mSystems. 2022 Dec 20;7(6):e0072322. doi: 10.1128/msystems.00723-22. Epub 2022 Oct 13. mSystems. 2022. PMID: 36226968 Free PMC article.
-
H-NS regulates the Vibrio parahaemolyticus type VI secretion system 1.Microbiology (Reading). 2014 Sep;160(Pt 9):1867-1873. doi: 10.1099/mic.0.080028-0. Epub 2014 Jul 1. Microbiology (Reading). 2014. PMID: 24987102 Free PMC article.
-
The Type VI secretion system: a versatile bacterial weapon.Microbiology (Reading). 2019 May;165(5):503-515. doi: 10.1099/mic.0.000789. Epub 2019 Mar 20. Microbiology (Reading). 2019. PMID: 30893029 Review.
-
Regulation of type VI secretion systems at the transcriptional, posttranscriptional and posttranslational level.Microbiology (Reading). 2023 Aug;169(8):001376. doi: 10.1099/mic.0.001376. Microbiology (Reading). 2023. PMID: 37552221 Free PMC article. Review.
Cited by
-
Role of the Type VI Secretion System in the Pathogenicity of Pseudomonas syringae pv. actinidiae, the Causative Agent of Kiwifruit Bacterial Canker.Front Microbiol. 2021 Feb 19;12:627785. doi: 10.3389/fmicb.2021.627785. eCollection 2021. Front Microbiol. 2021. PMID: 33679650 Free PMC article.
-
Adaptations of Vibrio parahaemolyticus to Stress During Environmental Survival, Host Colonization, and Infection.Front Microbiol. 2021 Oct 7;12:737299. doi: 10.3389/fmicb.2021.737299. eCollection 2021. Front Microbiol. 2021. PMID: 34690978 Free PMC article. Review.
-
A Rapid Fluorescence-Based Screen to Identify Regulators and Components of Interbacterial Competition Mechanisms in Bacteria.Methods Mol Biol. 2022;2427:11-24. doi: 10.1007/978-1-0716-1971-1_2. Methods Mol Biol. 2022. PMID: 35619021
-
Validation of the Thermo Scientific™ SureTect™ Vibrio cholerae, Vibrio parahaemolyticus, and Vibrio vulnificus PCR Assay for the Detection of Vibrio cholerae, Vibrio parahaemolyticus, and Vibrio vulnificus in Seafood Matrixes: AOAC Performance Tested MethodsSM 022301.J AOAC Int. 2023 Sep 1;106(5):1254-1277. doi: 10.1093/jaoacint/qsad061. J AOAC Int. 2023. PMID: 37243669 Free PMC article.
-
Genetic Basis of High-Pressure Tolerance of a Vibrio parahaemolyticus Mutant and Its Pathogenicity.Front Microbiol. 2022 Mar 31;13:827856. doi: 10.3389/fmicb.2022.827856. eCollection 2022. Front Microbiol. 2022. PMID: 35432286 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
