

Evidence that alternative transcriptional initiation is largely nonadaptive
- PMID: 30883542
- PMCID: PMC6438578
- DOI: 10.1371/journal.pbio.3000197
Evidence that alternative transcriptional initiation is largely nonadaptive
Abstract
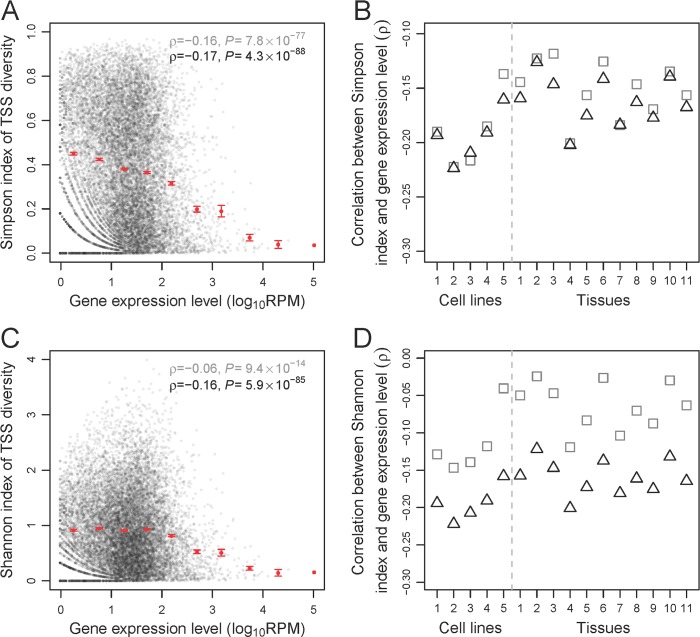
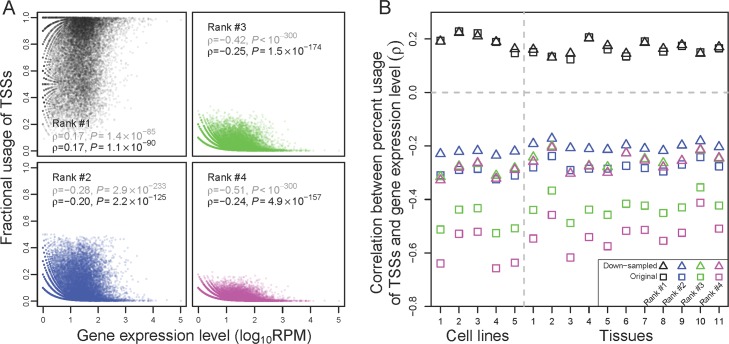
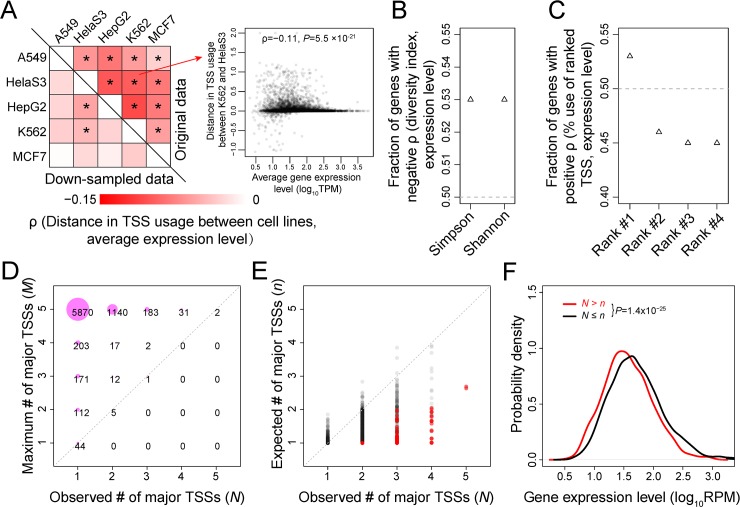
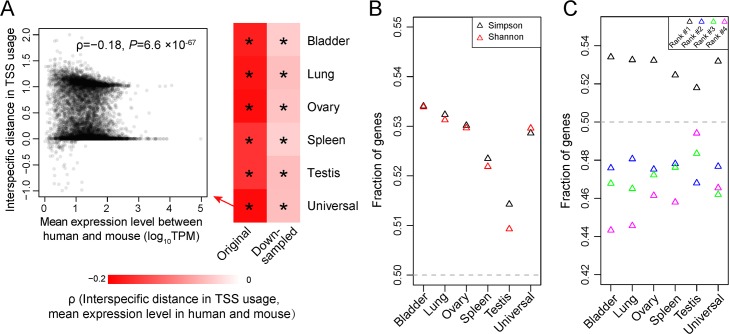
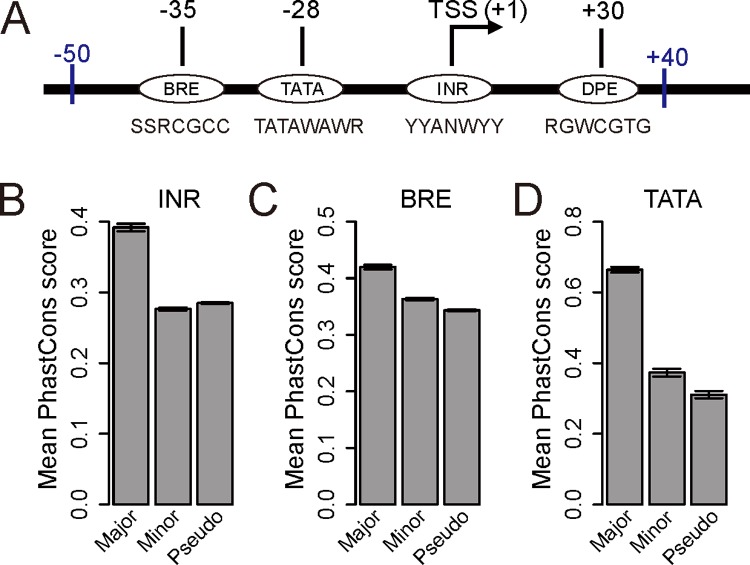
Alternative transcriptional initiation (ATI) refers to the frequent observation that one gene has multiple transcription start sites (TSSs). Although this phenomenon is thought to be adaptive, the specific advantage is rarely known. Here, we propose that each gene has one optimal TSS and that ATI arises primarily from imprecise transcriptional initiation that could be deleterious. This error hypothesis predicts that (i) the TSS diversity of a gene reduces with its expression level; (ii) the fractional use of the major TSS increases, but that of each minor TSS decreases, with the gene expression level; and (iii) cis-elements for major TSSs are selectively constrained, while those for minor TSSs are not. By contrast, the adaptive hypothesis does not make these predictions a priori. Our analysis of human and mouse transcriptomes confirms each of the three predictions. These and other findings strongly suggest that ATI predominantly results from molecular errors, requiring a major revision of our understanding of the precision and regulation of transcription.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
