

Centromere Repeats: Hidden Gems of the Genome
- PMID: 30884847
- PMCID: PMC6471113
- DOI: 10.3390/genes10030223
Centromere Repeats: Hidden Gems of the Genome
Abstract
Satellite DNAs are now regarded as powerful and active contributors to genomic and chromosomal evolution. Paired with mobile transposable elements, these repetitive sequences provide a dynamic mechanism through which novel karyotypic modifications and chromosomal rearrangements may occur. In this review, we discuss the regulatory activity of satellite DNA and their neighboring transposable elements in a chromosomal context with a particular emphasis on the integral role of both in centromere function. In addition, we discuss the varied mechanisms by which centromeric repeats have endured evolutionary processes, producing a novel, species-specific centromeric landscape despite sharing a ubiquitously conserved function. Finally, we highlight the role these repetitive elements play in the establishment and functionality of de novo centromeres and chromosomal breakpoints that underpin karyotypic variation. By emphasizing these unique activities of satellite DNAs and transposable elements, we hope to disparage the conventional exemplification of repetitive DNA in the historically-associated context of 'junk'.
Keywords: CENP-A; centromere drive; centromeric transcription; chromosome evolution; genetic conflict; repetitive DNA; satellite; transposable element.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
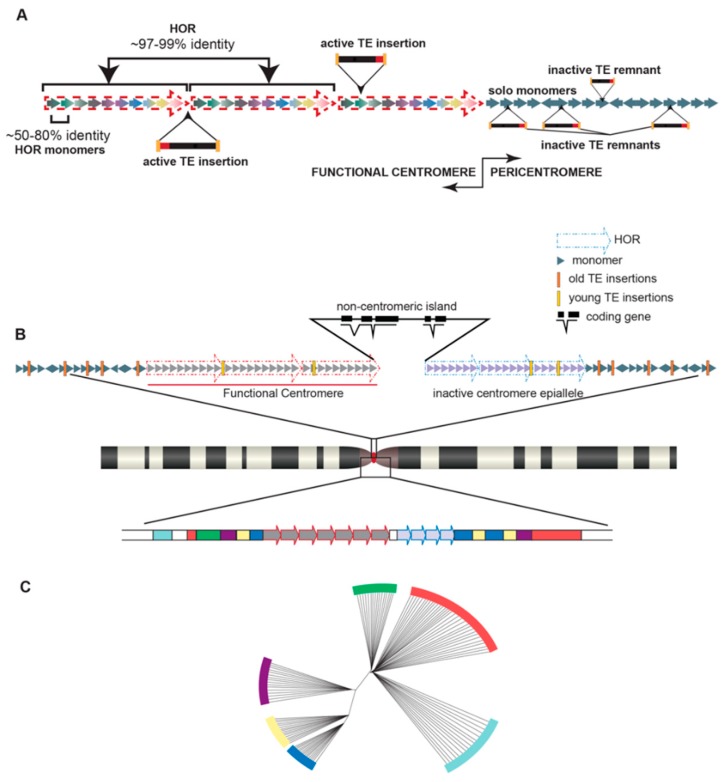
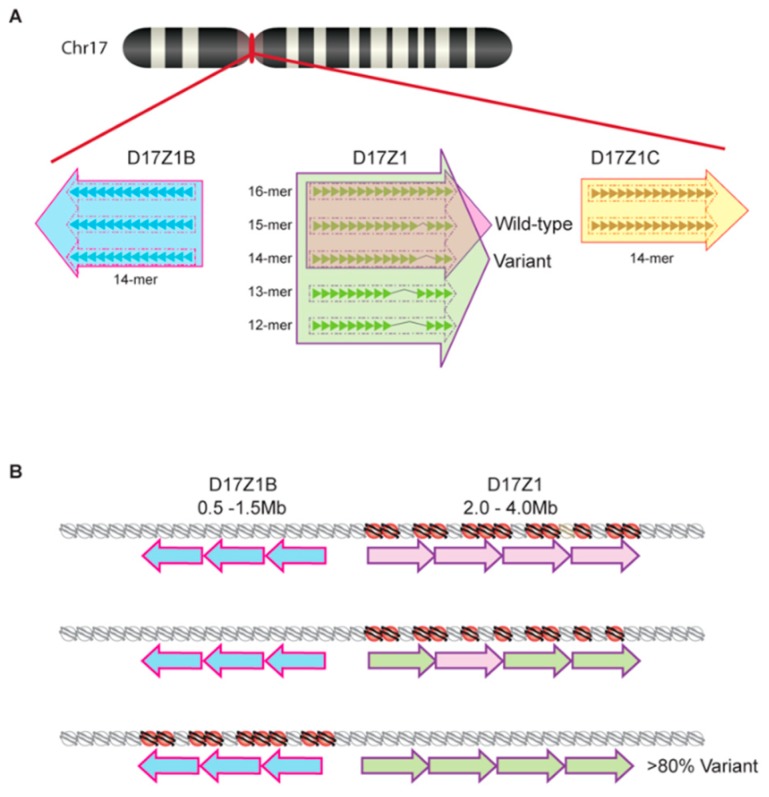
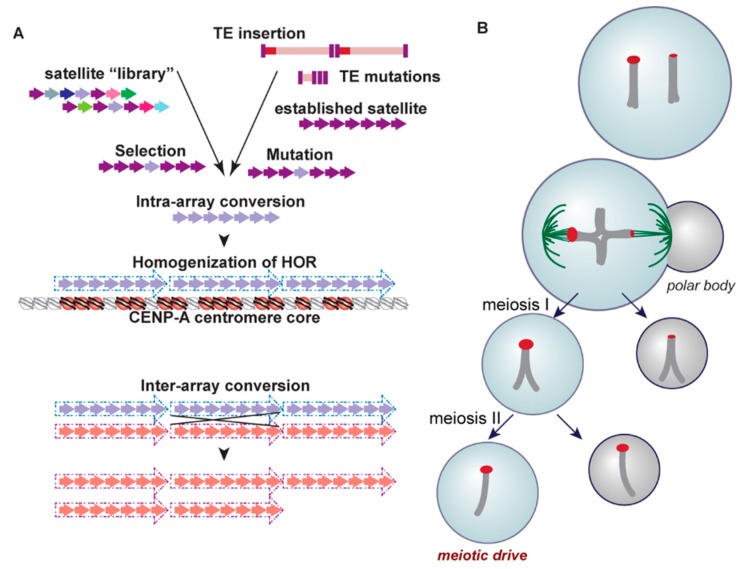
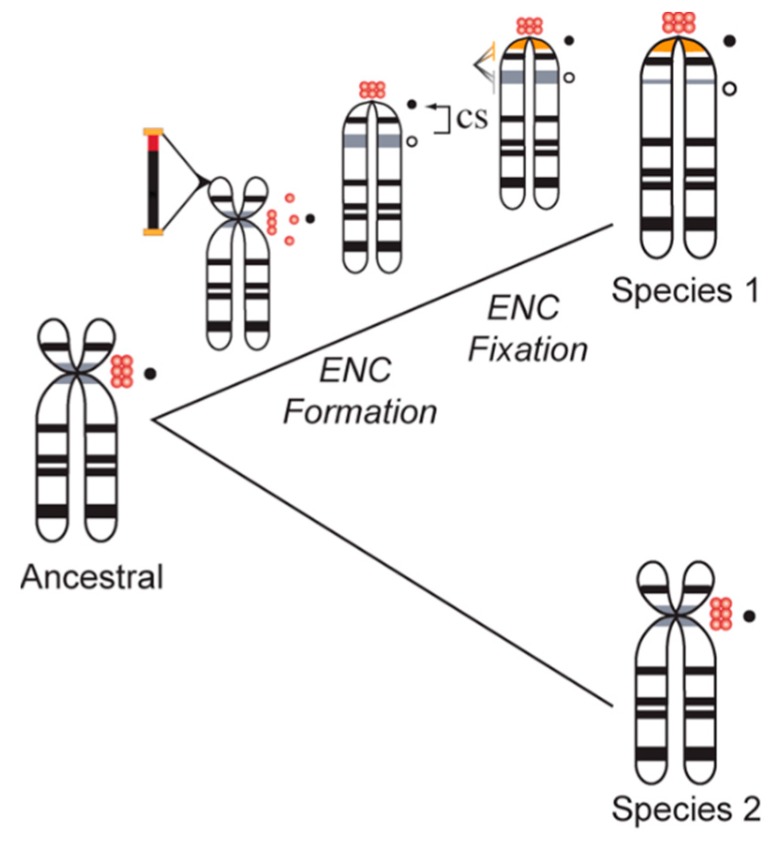
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
