

Protein arginine methyltransferase 5 (PRMT5) promotes survival of lymphoma cells via activation of WNT/β-catenin and AKT/GSK3β proliferative signaling
- PMID: 30885941
- PMCID: PMC6514641
- DOI: 10.1074/jbc.RA119.007640
Protein arginine methyltransferase 5 (PRMT5) promotes survival of lymphoma cells via activation of WNT/β-catenin and AKT/GSK3β proliferative signaling
Abstract
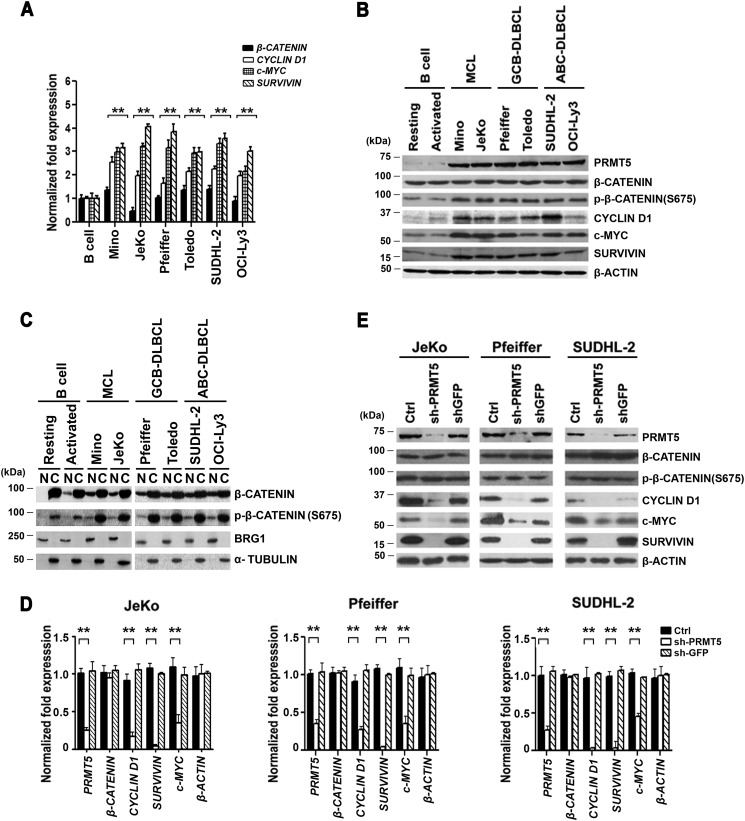
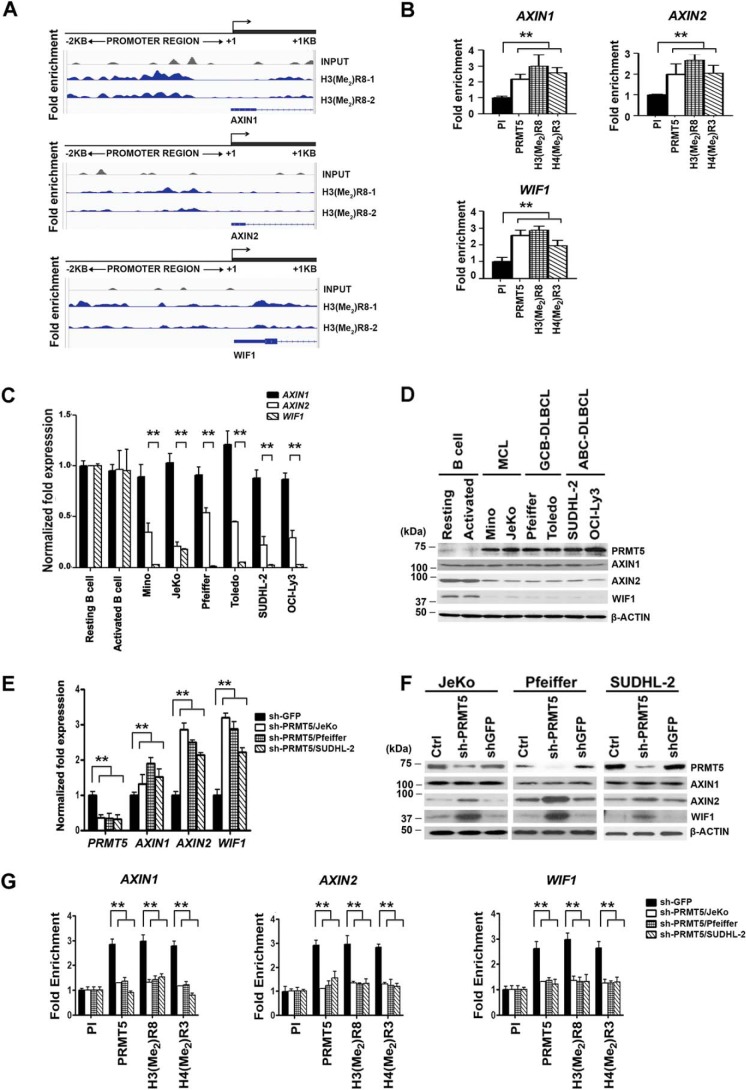
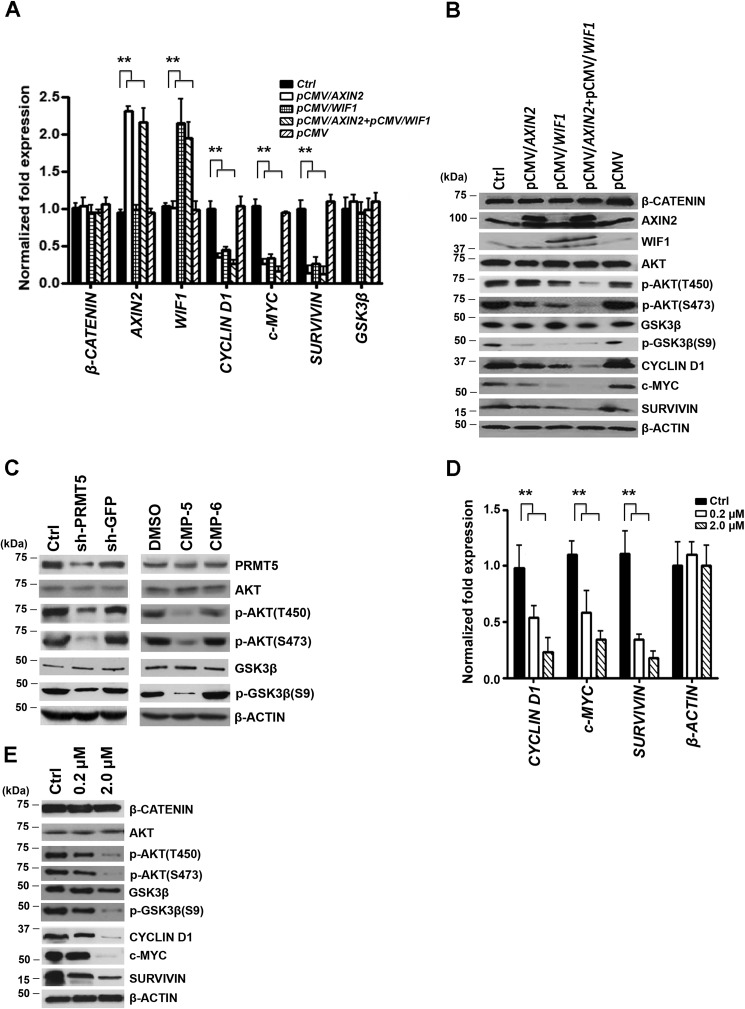
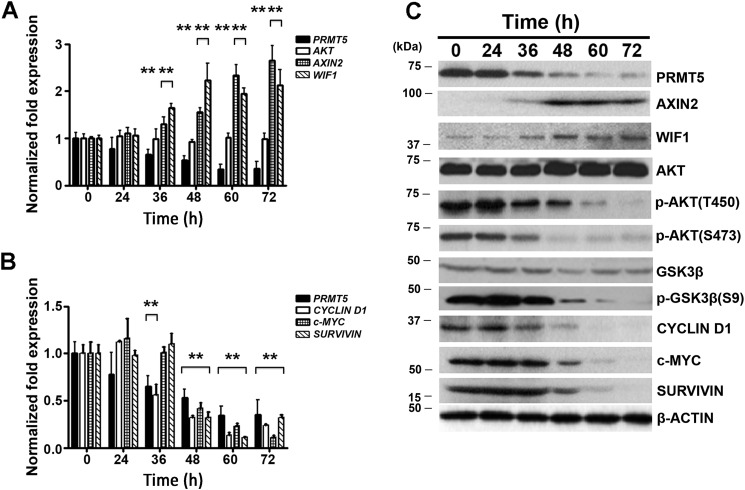
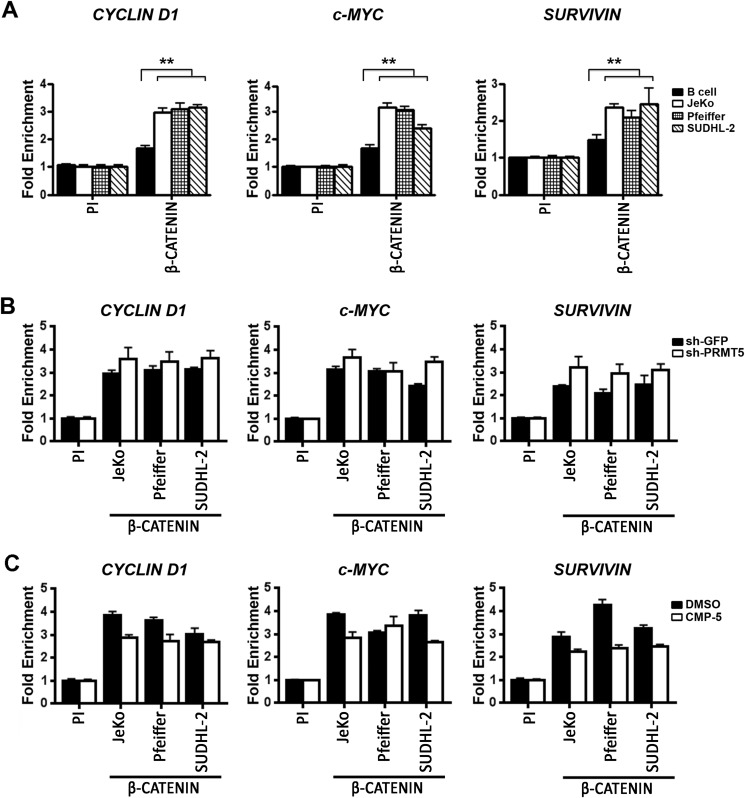
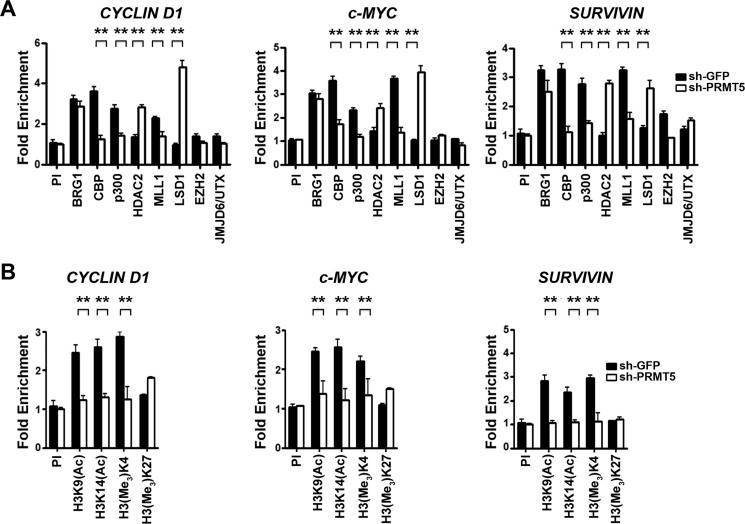
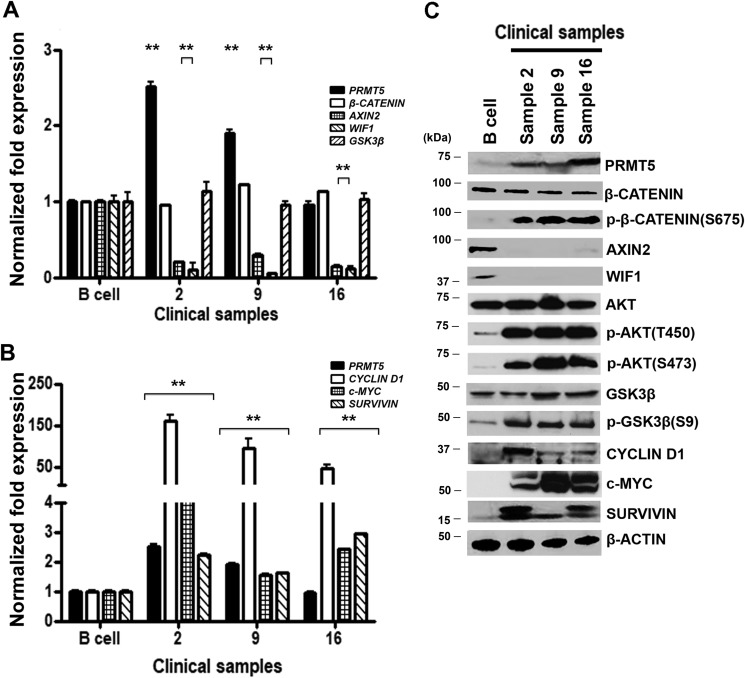
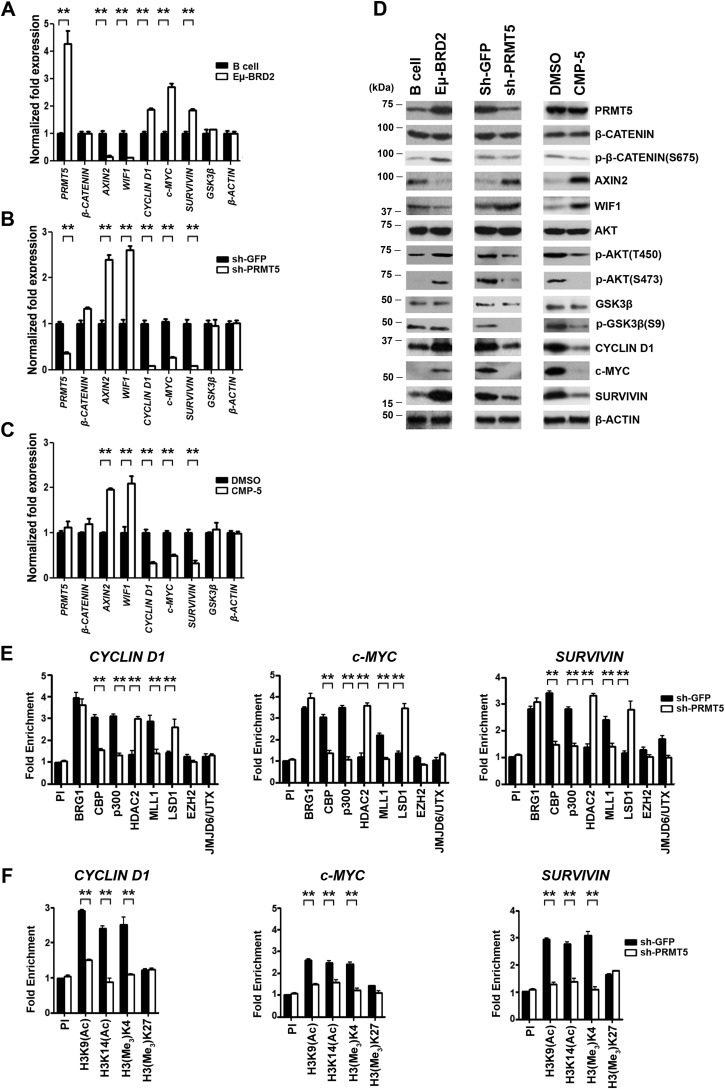
Epigenetic regulation by the type II protein arginine methyltransferase, PRMT5, plays an essential role in the control of cancer cell proliferation and tumorigenesis. In this report, we investigate the relationship between PRMT5 and WNT/β-CATENIN as well as AKT/GSK3β proliferative signaling in three different types of non-Hodgkin's lymphoma cell lines, clinical samples, and mouse primary lymphoma cells. We show that PRMT5 stimulates WNT/β-CATENIN signaling through direct epigenetic silencing of pathway antagonists, AXIN2 and WIF1, and indirect activation of AKT/GSK3β signaling. PRMT5 inhibition with either shRNA-mediated knockdown or a specific small molecule PRMT5 inhibitor, CMP-5, not only leads to derepression of WNT antagonists and decreased levels of active phospho-AKT (Thr-450 and Ser-473) and inactive phospho-GSK3β (Ser-9) but also results in decreased transcription of WNT/β-CATENIN target genes, CYCLIN D1, c-MYC, and SURVIVIN, and enhanced lymphoma cell death. Furthermore, PRMT5 inhibition leads to reduced recruitment of co-activators CBP, p300, and MLL1, as well as enhanced recruitment of co-repressors HDAC2 and LSD1 to the WNT/β-CATENIN target gene promoters. These results indicate that PRMT5 governs expression of prosurvival genes by promoting WNT/β-CATENIN and AKT/GSK3β proliferative signaling and that its inhibition induces lymphoma cell death, which warrants further clinical evaluation.
Keywords: Akt PKB; WNT antagonists; WNT/β-catenin signaling; Wnt signaling; histone; lymphoma; non-Hodgkin's lymphoma; protein arginine N-methyltransferase 5 (PRMT5); protein arginine methyltransferase 5 (PRMT5); protein kinase B (AKT)/glycogen synthase kinase 3β.
© 2019 Chung et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
Protein arginine methyltransferase 5 (PRMT5) activates WNT/β-catenin signalling in breast cancer cells via epigenetic silencing of DKK1 and DKK3.J Cell Mol Med. 2021 Feb;25(3):1583-1600. doi: 10.1111/jcmm.16260. Epub 2021 Jan 18. J Cell Mol Med. 2021. PMID: 33462997 Free PMC article.
-
Protein arginine methyltransferase 5 (PRMT5) inhibition induces lymphoma cell death through reactivation of the retinoblastoma tumor suppressor pathway and polycomb repressor complex 2 (PRC2) silencing.J Biol Chem. 2013 Dec 6;288(49):35534-47. doi: 10.1074/jbc.M113.510669. Epub 2013 Nov 4. J Biol Chem. 2013. PMID: 24189068 Free PMC article.
-
PRMT5 Promotes Human Lung Cancer Cell Apoptosis via Akt/Gsk3β Signaling Induced by Resveratrol.Cell Transplant. 2019 Dec;28(12):1664-1673. doi: 10.1177/0963689719885083. Epub 2019 Oct 30. Cell Transplant. 2019. PMID: 31665911 Free PMC article.
-
Aberrant miRNA expression and protein arginine methyltransferase 5 (PRMT5) in cancer: A review.Biochim Biophys Acta Mol Cell Res. 2025 Mar;1872(3):119923. doi: 10.1016/j.bbamcr.2025.119923. Epub 2025 Feb 22. Biochim Biophys Acta Mol Cell Res. 2025. PMID: 39993609 Review.
-
Cadmium inhibits neural stem/progenitor cells proliferation via MitoROS-dependent AKT/GSK-3β/β-catenin signaling pathway.J Appl Toxicol. 2021 Dec;41(12):1998-2010. doi: 10.1002/jat.4179. Epub 2021 May 11. J Appl Toxicol. 2021. PMID: 33977565 Review.
Cited by
-
PRMT5 inhibition drives therapeutic vulnerability to combination treatment with BCL-2 inhibition in mantle cell lymphoma.Blood Adv. 2023 Oct 24;7(20):6211-6224. doi: 10.1182/bloodadvances.2023009906. Blood Adv. 2023. PMID: 37327122 Free PMC article.
-
TP53 mutations and RNA-binding protein MUSASHI-2 drive resistance to PRMT5-targeted therapy in B-cell lymphoma.Nat Commun. 2022 Sep 27;13(1):5676. doi: 10.1038/s41467-022-33137-8. Nat Commun. 2022. PMID: 36167829 Free PMC article.
-
Targeting Epigenetic Modifiers of Tumor Plasticity and Cancer Stem Cell Behavior.Cells. 2022 Apr 21;11(9):1403. doi: 10.3390/cells11091403. Cells. 2022. PMID: 35563709 Free PMC article. Review.
-
Resistance to PRMT5-targeted therapy in mantle cell lymphoma.Blood Adv. 2024 Jan 9;8(1):150-163. doi: 10.1182/bloodadvances.2023010554. Blood Adv. 2024. PMID: 37782774 Free PMC article.
-
Protein arginine methyltransferase 5 (PRMT5) activates WNT/β-catenin signalling in breast cancer cells via epigenetic silencing of DKK1 and DKK3.J Cell Mol Med. 2021 Feb;25(3):1583-1600. doi: 10.1111/jcmm.16260. Epub 2021 Jan 18. J Cell Mol Med. 2021. PMID: 33462997 Free PMC article.
References
-
- Swerdlow S. H., Campo E., Seto M., and Müller-Hemrelink H. K. (2008) in WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues (Swerdlow S. H., Campo E., Harris N. L., Jaffe E. S., Pileri S. A., Stein H., Thiele J., and Vardiman J. W., eds) pp. 229–232. IARC Publishing Group, Lyon, France
-
- Alizadeh A. A., Eisen M. B., Davis R. E., Ma C., Lossos I. S., Rosenwald A., Boldrick J. C., Sabet H., Tran T., Yu X., Powell J. I., Yang L., Marti G. E., Moore T., Hudson J. Jr., et al. (2000) Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature 403, 503–511 10.1038/35000501 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
