

Antibody responses to viral infections: a structural perspective across three different enveloped viruses
- PMID: 30886356
- PMCID: PMC6818971
- DOI: 10.1038/s41564-019-0392-y
Antibody responses to viral infections: a structural perspective across three different enveloped viruses
Abstract
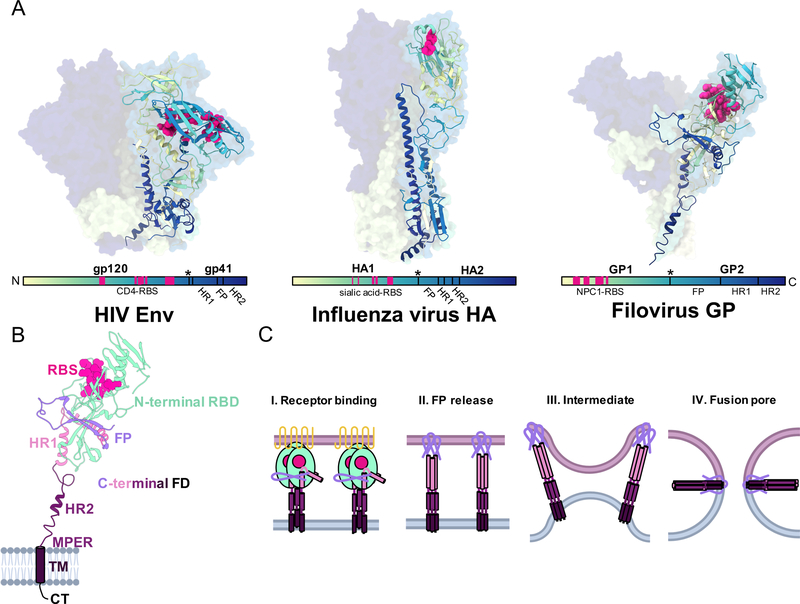
Antibodies serve as critical barriers to viral infection. Humoral immunity to a virus is achieved through the dual role of antibodies in communicating the presence of invading pathogens in infected cells to effector cells, and in interfering with processes essential to the viral life cycle (chiefly entry into the host cell). For individuals that successfully control infection, virus-elicited antibodies can provide lifelong surveillance and protection from future insults. One approach to understand the nature of a successful immune response has been to utilize structural biology to uncover the molecular details of antibodies derived from vaccines or natural infection and how they interact with their cognate microbial antigens. The ability to isolate antigen-specific B-cells and rapidly solve structures of functional, monoclonal antibodies in complex with viral glycoprotein surface antigens has greatly expanded our knowledge of the sites of vulnerability on viruses. In this Review, we compare the adaptive humoral immune responses to human immunodeficiency virus (HIV), influenza and filoviruses, with a particular focus on neutralizing antibodies. The pathogenesis of each of these viruses is quite different, providing an opportunity for comparison of immune responses: HIV causes a persistent, chronic infection; influenza, an acute infection with multiple exposures during a lifetime and annual vaccination; filoviruses, a virulent, acute infection. Neutralizing antibodies that develop under these different constraints are therefore sentinels that can provide insight into the underlying humoral immune responses, as well as important lessons to guide future development of vaccines and immunotherapeutics.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
