

RNA structure maps across mammalian cellular compartments
- PMID: 30886404
- PMCID: PMC6640855
- DOI: 10.1038/s41594-019-0200-7
RNA structure maps across mammalian cellular compartments
Abstract
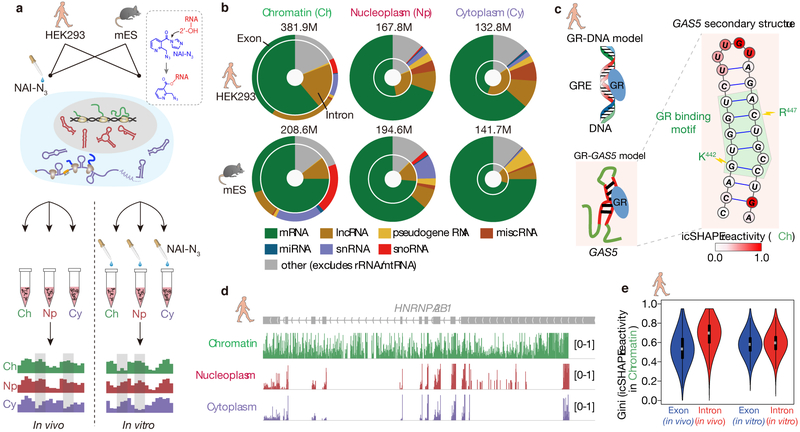
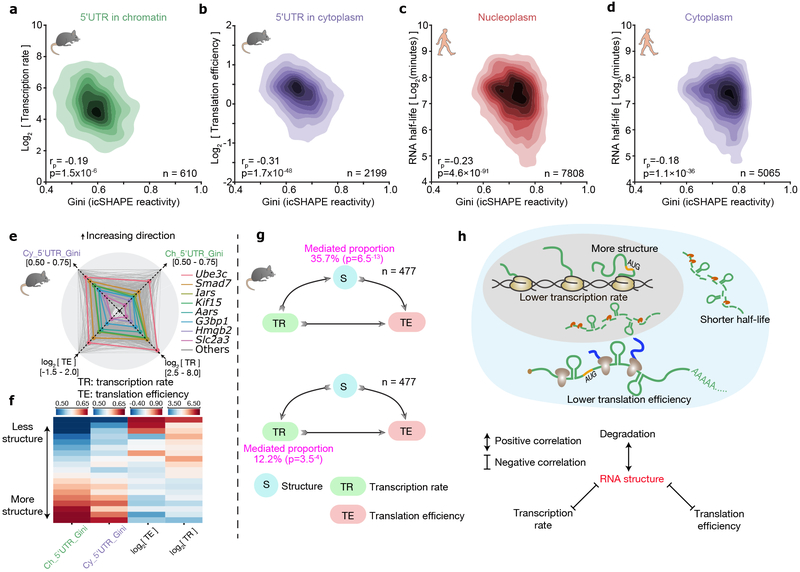
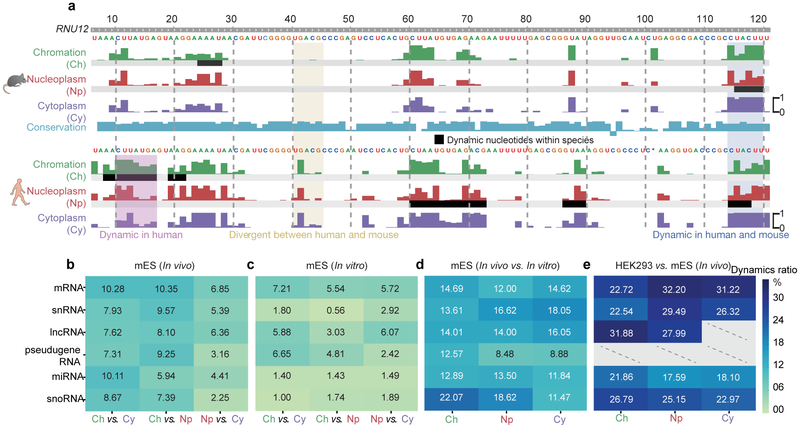
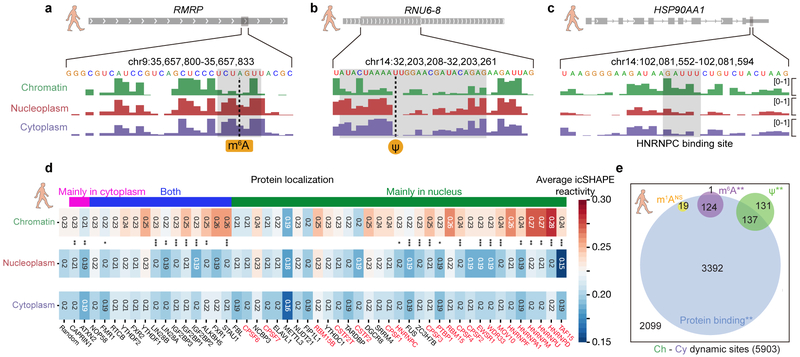
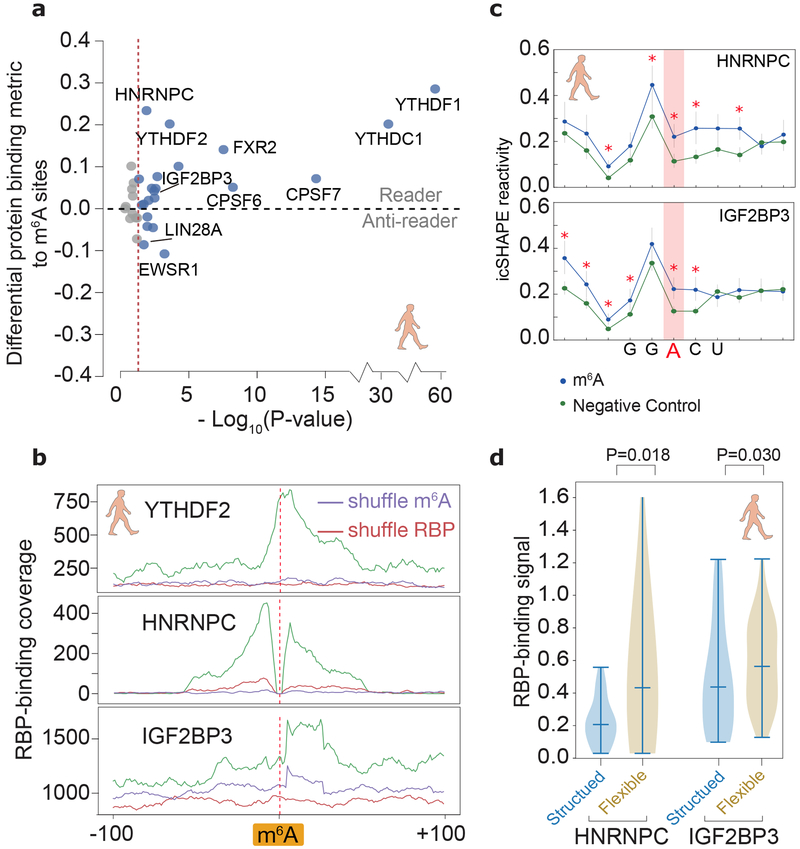
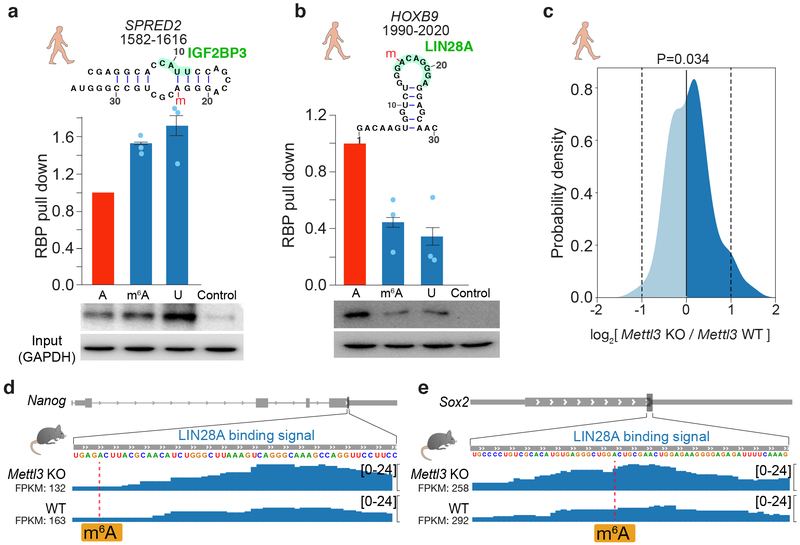
RNA structure is intimately connected to each step of gene expression. Recent advances have enabled transcriptome-wide maps of RNA secondary structure, called 'RNA structuromes'. However, previous whole-cell analyses lacked the resolution to unravel the landscape and also the regulatory mechanisms of RNA structural changes across subcellular compartments. Here we reveal the RNA structuromes in three compartments, chromatin, nucleoplasm and cytoplasm, in human and mouse cells. The cytotopic structuromes substantially expand RNA structural information and enable detailed investigation of the central role of RNA structure in linking transcription, translation and RNA decay. We develop a resource with which to visualize the interplay of RNA-protein interactions, RNA modifications and RNA structure and predict both direct and indirect reader proteins of RNA modifications. We also validate a novel role for the RNA-binding protein LIN28A as an N6-methyladenosine modification 'anti-reader'. Our results highlight the dynamic nature of RNA structures and its functional importance in gene regulation.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
