

Development of drug-inducible CRISPR-Cas9 systems for large-scale functional screening
- PMID: 30890156
- PMCID: PMC6425629
- DOI: 10.1186/s12864-019-5601-9
Development of drug-inducible CRISPR-Cas9 systems for large-scale functional screening
Abstract
Background: Large-scale genetic screening using CRISPR-Cas9 technology has emerged as a powerful approach to uncover and validate gene functions. The ability to control the timing of genetic perturbation during CRISPR screens will facilitate precise dissection of dynamic and complex biological processes. Here, we report the optimization of a drug-inducible CRISPR-Cas9 system that allows high-throughput gene interrogation with a temporal control.
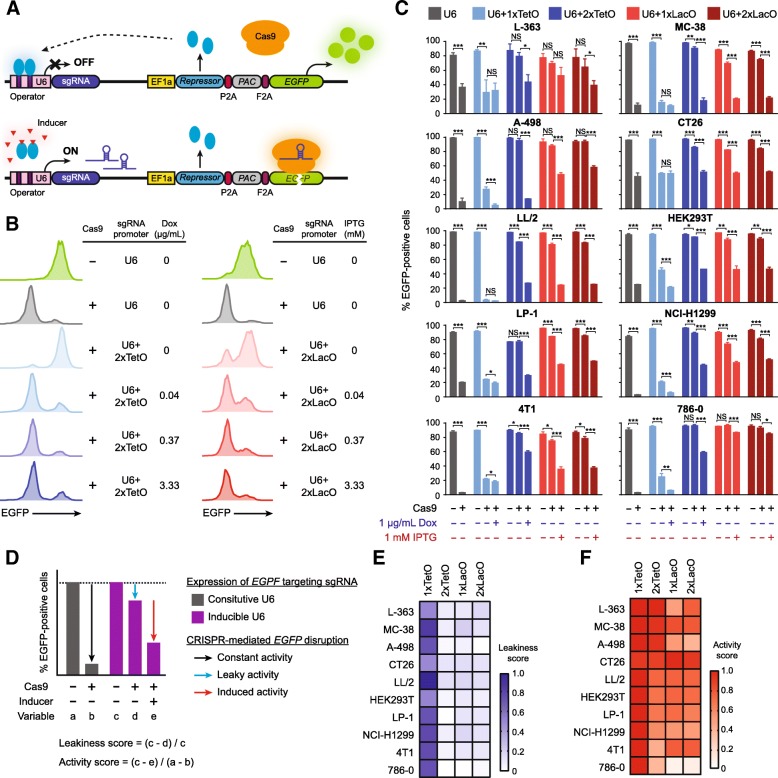
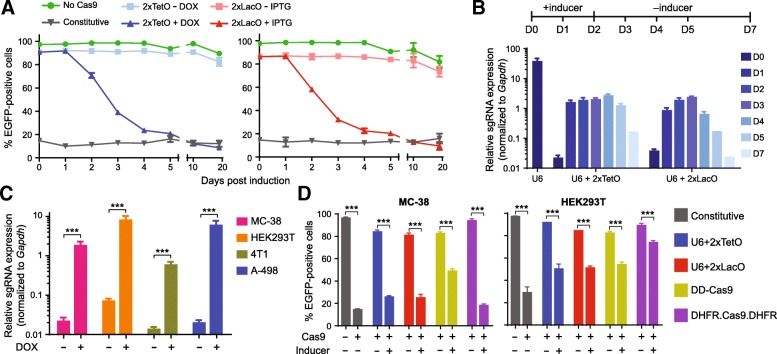
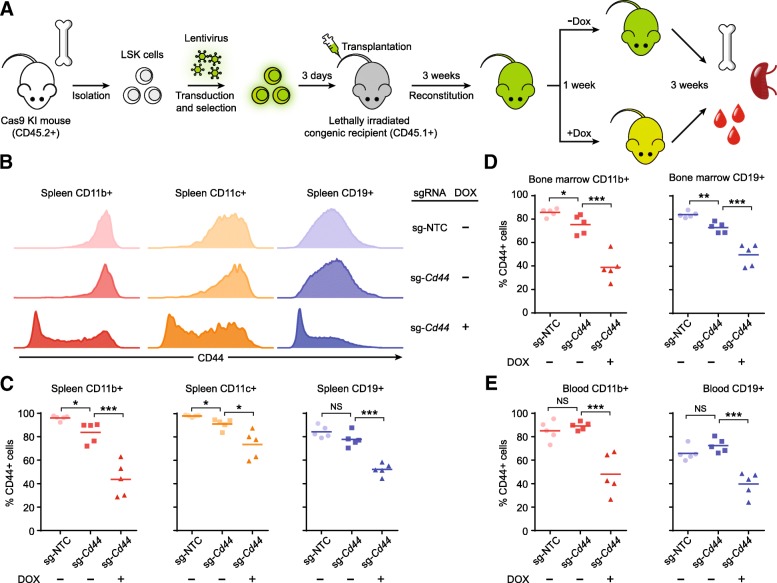
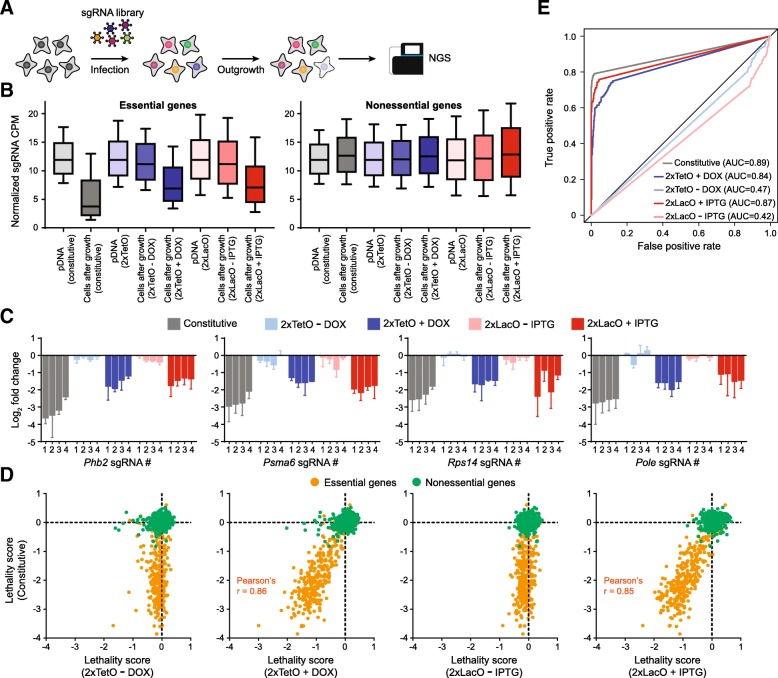
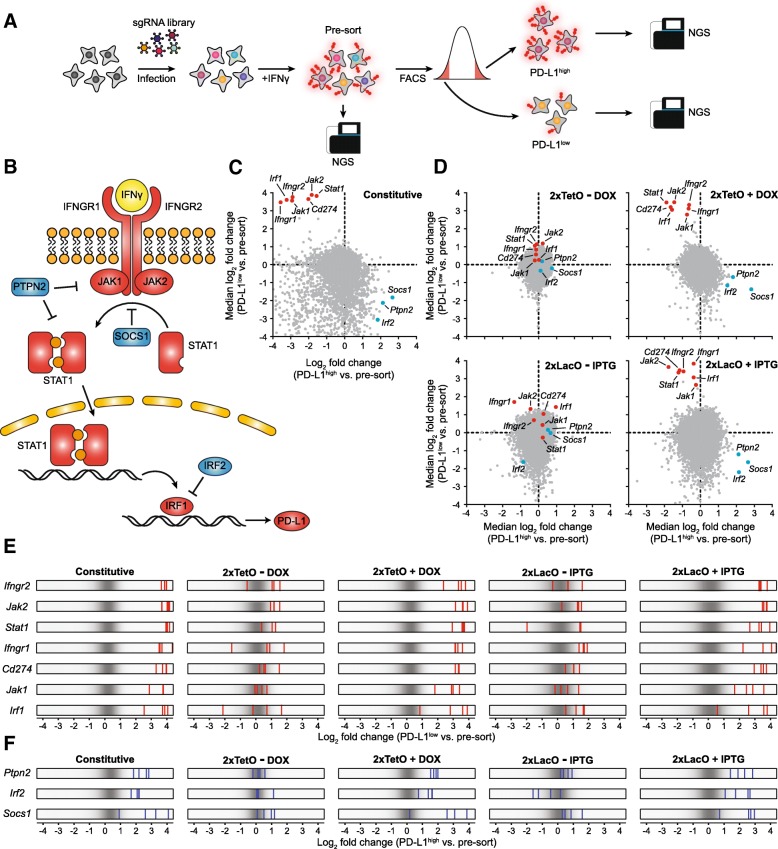
Results: We designed multiple drug-inducible sgRNA expression vectors and measured their activities using an EGFP gene disruption assay in 11 human and mouse cell lines. The optimal design allows for a tight and inducible control of gene knockout in vitro, and in vivo during a seven-week-long experiment following hematopoietic reconstitution in mice. We next performed parallel genome-wide loss-of-function screens using the inducible and constitutive CRISPR-Cas9 systems. In proliferation-based dropout screens, these two approaches have similar performance in discriminating essential and nonessential genes. In a more challenging phenotypic assay that requires cytokine stimulation and cell staining, we observed similar sensitivity of the constitutive and drug-induced screening approaches in detecting known hits. Importantly, we demonstrate minimal leakiness of our inducible CRISPR screening platforms in the absence of chemical inducers in large-scale settings.
Conclusions: In this study, we have developed a drug-inducible CRISPR-Cas9 system that shows high cleavage efficiency upon induction but low background activity. Using this system, we have achieved inducible gene disruption in a wide range of cell types both in vitro and in vivo. For the first time, we present a systematic side-by-side comparison of constitutive and drug-inducible CRISPR-Cas9 platforms in large-scale functional screens. We demonstrate the tightness and efficiency of our drug-inducible CRISPR-Cas9 system in genome-wide pooled screening. Our design increases the versatility of CRISPR-based genetic screening and represents a significant upgrade on existing functional genomics toolbox.
Keywords: CRISPR; Functional genomics; Gene editing.
Conflict of interest statement
Ethics approval and consent to participate
All studies involving animals were performed according to protocols reviewed and approved by the Abbvie IACUC.
Competing interests
All authors were employees of AbbVie at the time of the study.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
