

Pancreatic cancer-initiating cell exosome message transfer into noncancer-initiating cells: the importance of CD44v6 in reprogramming
- PMID: 30890157
- PMCID: PMC6425561
- DOI: 10.1186/s13046-019-1129-8
Pancreatic cancer-initiating cell exosome message transfer into noncancer-initiating cells: the importance of CD44v6 in reprogramming
Erratum in
-
Correction: Pancreatic cancer-initiating cell exosome message transfer into noncancer-initiating cells: the importance of CD44v6 in reprogramming.J Exp Clin Cancer Res. 2024 Oct 31;43(1):293. doi: 10.1186/s13046-024-03216-3. J Exp Clin Cancer Res. 2024. PMID: 39478595 Free PMC article. No abstract available.
Retraction in
-
Retraction Note: Pancreatic cancer-initiating cell exosome message transfer into noncancer-initiating cells: the importance of CD44v6 in reprogramming.J Exp Clin Cancer Res. 2025 Feb 28;44(1):78. doi: 10.1186/s13046-025-03339-1. J Exp Clin Cancer Res. 2025. PMID: 40022185 Free PMC article. No abstract available.
Abstract
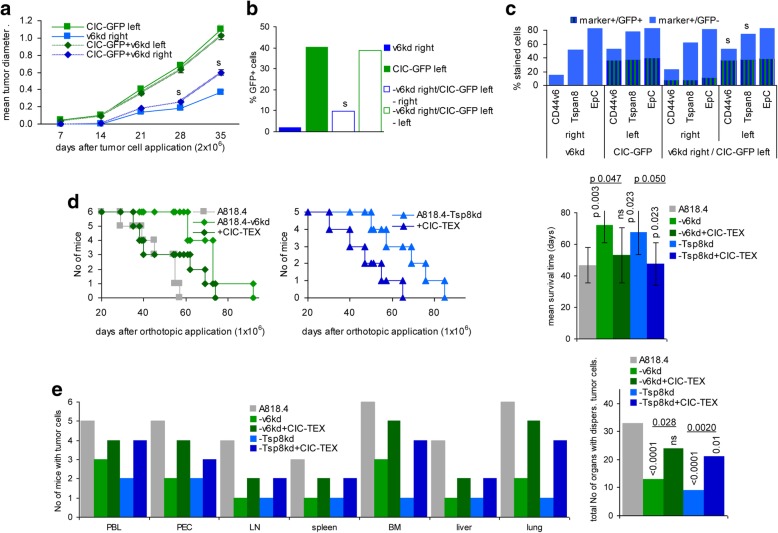
Background: Cancer-initiating cell (CIC) exosomes (CIC-TEX) are suggested reprogramming Non-CIC. Mode of message transfer and engagement of CIC-markers being disputed, we elaborated the impact of CD44v6 and Tspan8 on the response of Non-CIC.
Methods: Non-metastasizing CD44v6- and Tspan8-knockdown (kd) pancreatic cancer cells served as Non-CIC. CIC-TEX coculture-induced changes were evaluated by deep-sequencing and functional assays. Tumor progression was surveyed during in vivo CIC-TEX treatment.
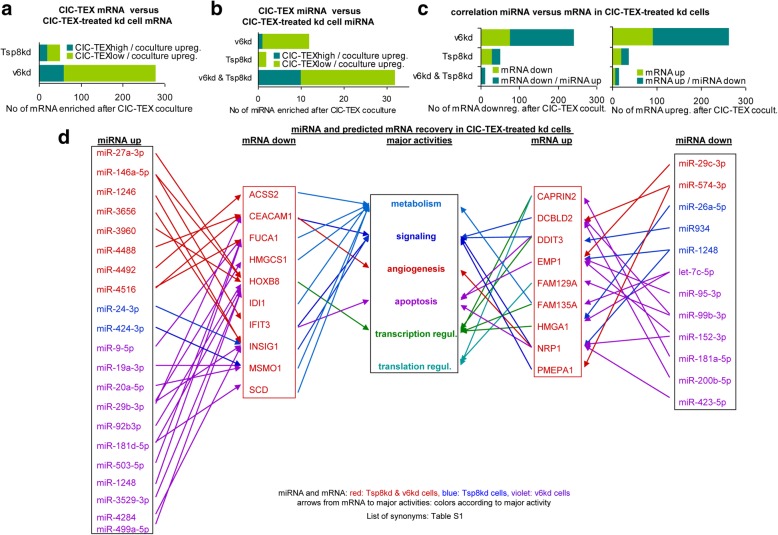
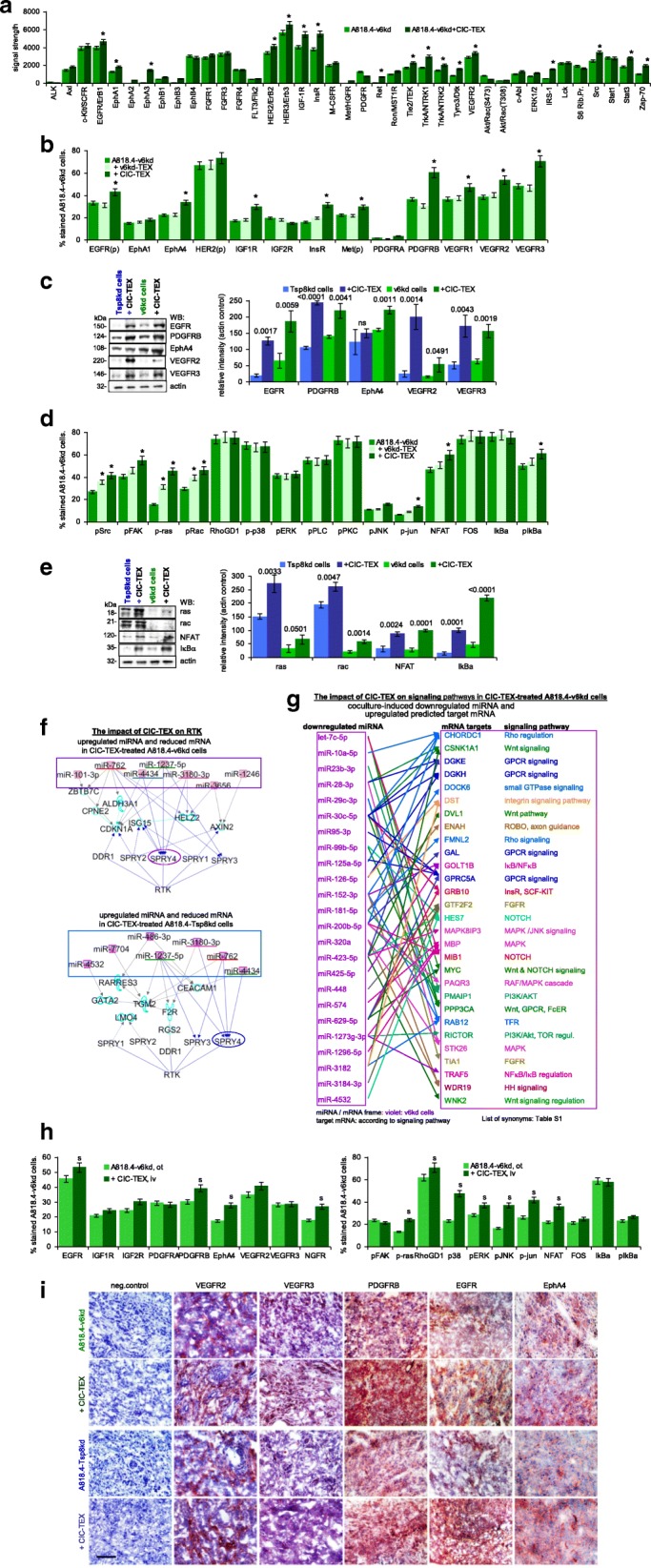
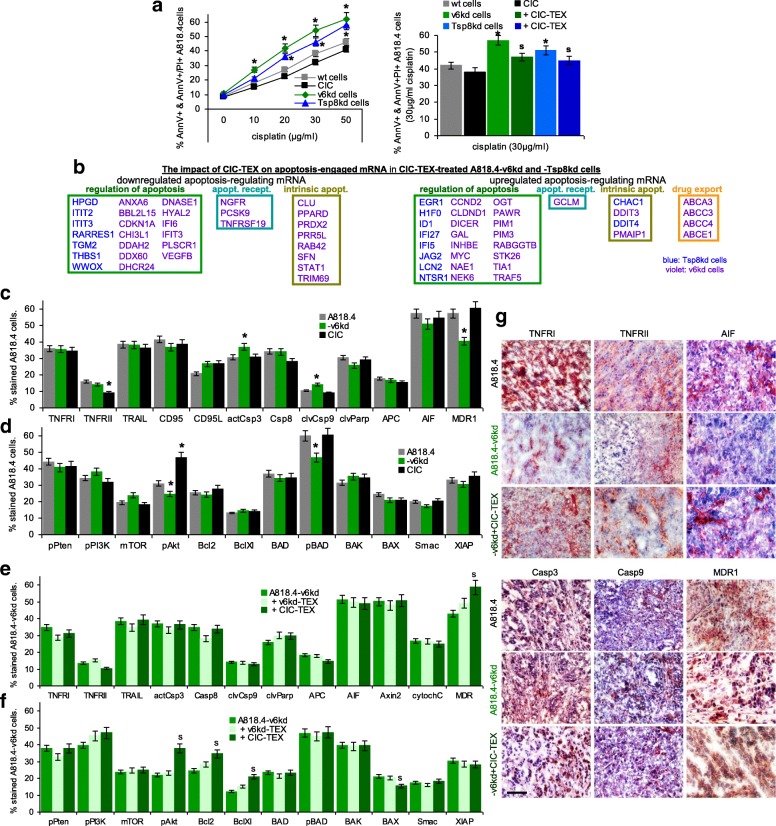
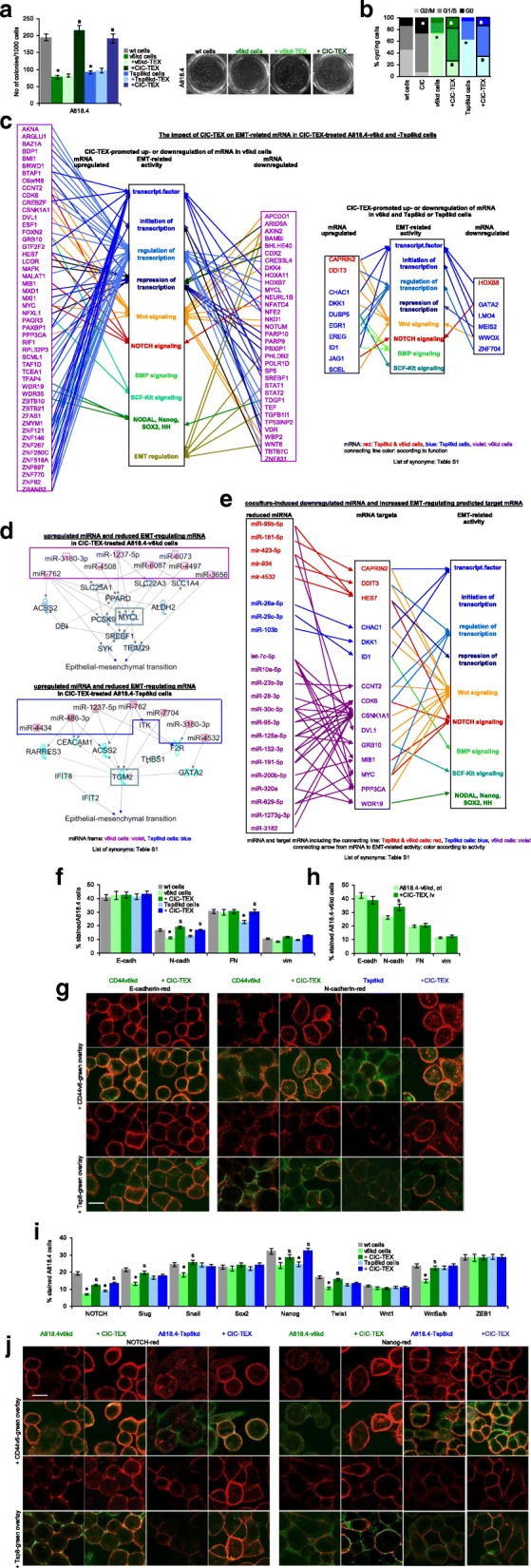
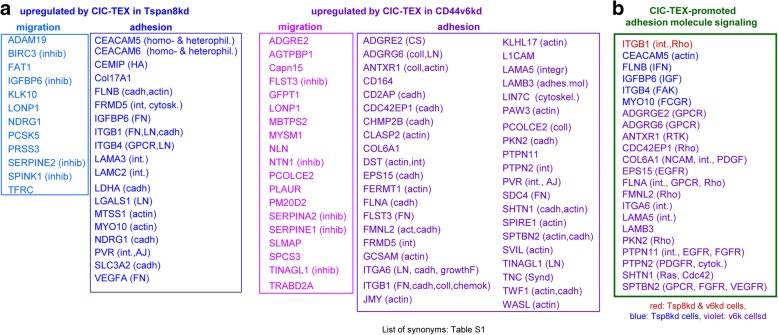
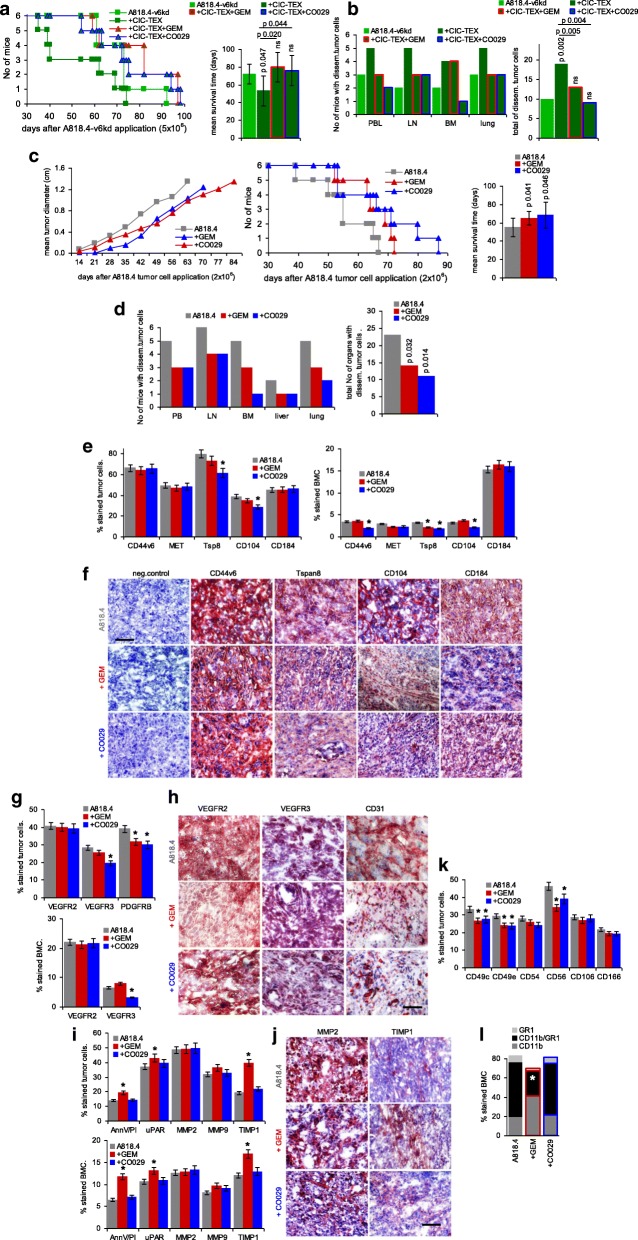
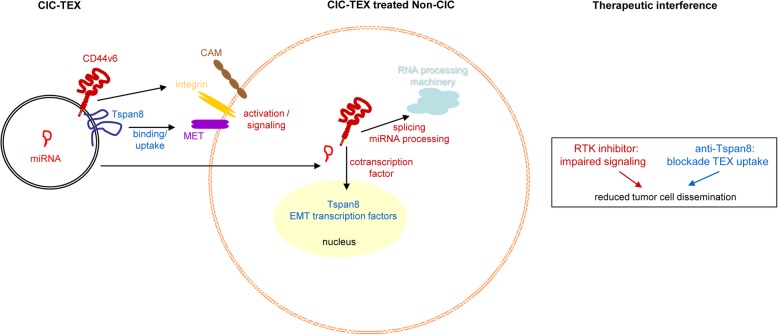
Results: Deep-sequencing of CIC-TEX-cocultured CD44v6kd-Non-CIC revealed pronounced mRNA changes in signaling, transport, transcription and translation; altered miRNA affected metabolism, signaling and transcription. CIC-TEX coculture-induced changes in Tspan8kd-Non-CIC mostly relied on CIC-TEX-Tspan8 being required for targeting. CIC-TEX transfer supported apoptosis resistance and significantly promoted epithelial mesenchymal transition, migration, invasion and (lymph)angiogenesis of the kd Non-CIC in vitro and in vivo, deep-sequencing allowing individual mRNA and miRNA assignment to altered functions. Importantly, CIC-TEX act as a hub, initiated by CD44v6-dependent RTK, GPCR and integrin activation and involving CD44v6-assisted transcription and RNA processing. Accordingly, a kinase inhibitor hampered CIC-TEX-fostered tumor progression, which was backed by an anti-Tspan8 blockade of CIC-TEX binding.
Conclusions: This in depth report on the in vitro and in vivo impact of CIC-TEX on CD44v6kd and Tspan8kd Non-CIC unravels hub CIC-TEX activity, highlighting a prominent contribution of the CIC-markers CD44v6 to signaling cascade activation, transcription, translation and miRNA processing in Non-CIC and of Tspan8 to CIC-TEX targeting. Blocking CIC-TEX binding/uptake and uptake-initiated target cell activation significantly mitigated the deleterious CIC-TEX impact on CD44v6kd and Tspan8kd Non-CIC.
Keywords: CD44v6; Exosome biogenesis; Exosome message transfer; Non-cancer stem cell reprogramming; Pancreatic cancer stem cells; Tspan8.
Conflict of interest statement
Authors’ information
The manuscript was edited for proper English language, grammar, punctuation, spelling, and overall style by one or more of the highly qualified native English speaking editors at Wiley Editing Services.
Ethics approval
Animal experiments were approved by the Government of Baden-Wuerttemberg, Germany. Animals were housed under specific pathogen-free conditions at the central animal facility of the University of Heidelberg (IBF) (Interfakultäre Biomedizinische Forschungseinrichtung). Yearly reports on animal experiments were delivered to the Government of Baden-Wuerttemberg, Germany, according to the rules of the European Union Directive 2010/63/EU for animal experiments.
Consent for publication
not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
