

Impact of constitutional TET2 haploinsufficiency on molecular and clinical phenotype in humans
- PMID: 30890702
- PMCID: PMC6424975
- DOI: 10.1038/s41467-019-09198-7
Impact of constitutional TET2 haploinsufficiency on molecular and clinical phenotype in humans
Abstract
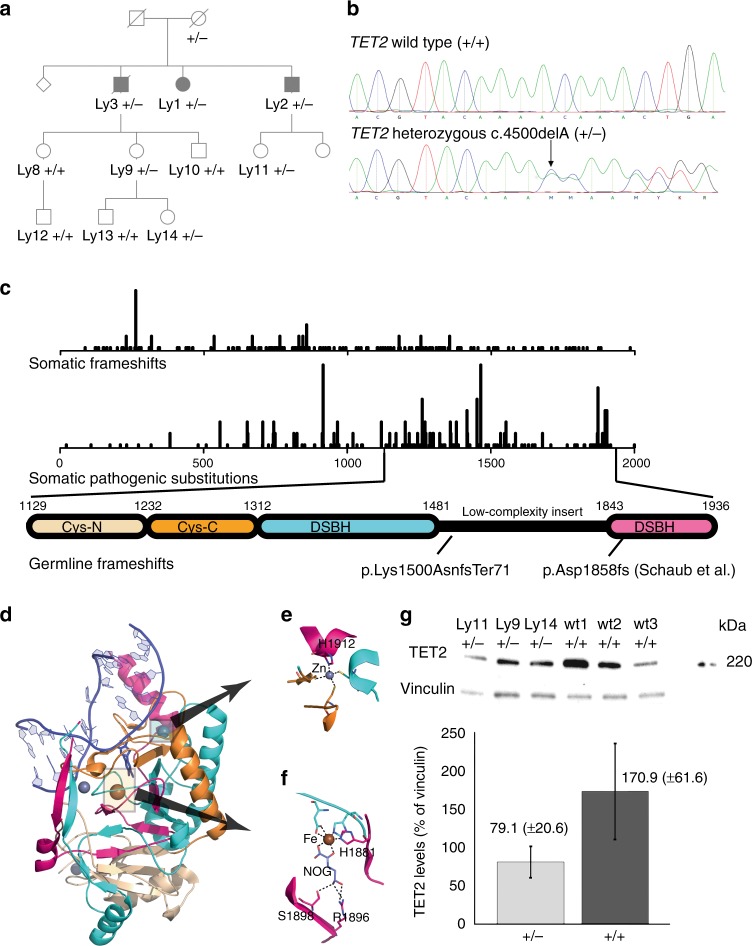
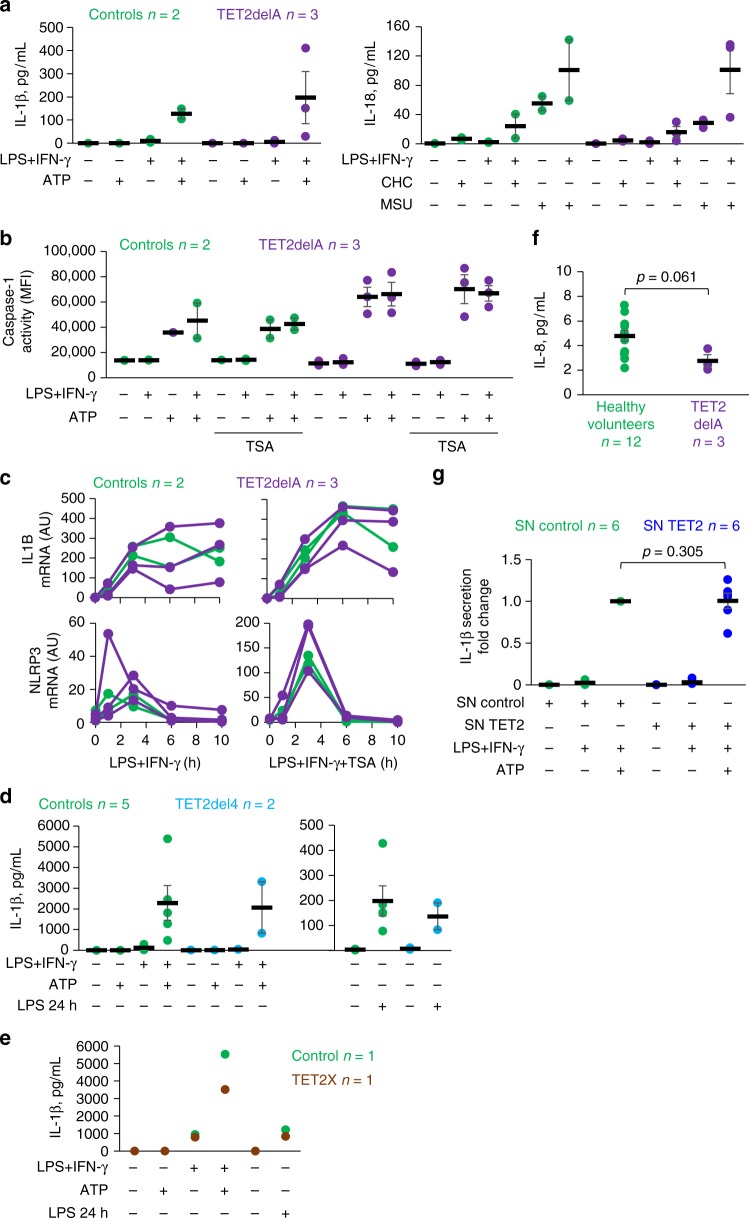
Clonal hematopoiesis driven by somatic heterozygous TET2 loss is linked to malignant degeneration via consequent aberrant DNA methylation, and possibly to cardiovascular disease via increased cytokine and chemokine expression as reported in mice. Here, we discover a germline TET2 mutation in a lymphoma family. We observe neither unusual predisposition to atherosclerosis nor abnormal pro-inflammatory cytokine or chemokine expression. The latter finding is confirmed in cells from three additional unrelated TET2 germline mutation carriers. The TET2 defect elevates blood DNA methylation levels, especially at active enhancers and cell-type specific regulatory regions with binding sequences of master transcription factors involved in hematopoiesis. The regions display reduced methylation relative to all open chromatin regions in four DNMT3A germline mutation carriers, potentially due to TET2-mediated oxidation. Our findings provide insight into the interplay between epigenetic modulators and transcription factor activity in hematological neoplasia, but do not confirm the putative role of TET2 in atherosclerosis.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Keaney, J. F. CHIP-ping away at atherosclerosis. N. Engl. J. Med. 10.1056/NEJMe1706173 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
